





(C) 2012 Daniel O. Wcislo. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Orchid bees (Euglossini) are a diverse and widespread Neotropical taxon comprised of five genera and nearly 200 species (
Aerial nests of Euglossa are either built on the undersides of leaves, or are top-shaped and attached to plant stems (e.g.,
We studied Euglossa hyacinthina at the species’ type locality in cloud forest near Fortuna, Panamá (Chiriqui Province) (elevation, ~ 1400 m). Nests were relatively abundant on small composites, mainly Baccharis pedunculata, between the Edwin Fabrega Hydroelectric Dam and about 2 km S of the Smithsonian Tropical Research Institute’s Jorge L. Arauz Center for Tropical Investigations, along the side of the road running from Los Planes to Chiriqui Grande.
Nest construction behavior was recorded with a GoPro HD Hero2 video camera mounted on a support approximately 25 cm from the nest. A partially constructed nest was discovered on 30 August 2012, and video recordings were made from approximately 9:00 AM to 12:00 PM and 2:00 PM to 4:30 PM on 31 August, with additional photos taken on 2 September. Additional observations on cell construction were made intermittently between May 2011 and January 2012 from nests in which viewing windows were cut into the envelope. Estimates of the amount of work completed were taken from digital photos. Nests were collected very late in the afternoon, presumably when most bees were home. Nest measurements were made with a Mitutoyo digital caliper. Means are given + one standard deviation. Statistical tests were done using PASW Statistics v 18.0. A voucher specimen is deposited in the Museo de los Invertebrados, Universidad de Panamá, and the STRI Dry Reference Collection.
Results and discussionTwo of 18 nests were presumably recently constructed, as inferred from the light orange color of the resin and the relatively pliable wall of the envelopes, and most (17) were relatively young, in contrast to an older nest that was dark brown, and brittle, as are abandoned nests (also
When first discovered, the nest envelope was approximately 1/3 completed (Figure 1A) and grew slowly each day (Figure 1B). Only a single female worked on constructing the new nest. This female made repeated foraging trips for resin lasting from 7 min 43 sec to 14 min 19 sec (x– = 619 + 138 sec, N = 7), returning to the nest with large masses of resin on the hind corbiculae, which she deposited in a small pile on the inner surface of the envelope as a temporary cache. These trips were faster than resin foraging in other Euglossa [means of 24.3 min and 18.2 min, respectively, for Euglossa atroveneta (
During construction the bee walked to the cache of resin and removed a small piece with her mandibles, and then went to the edge of the nest envelope, where she applied the resin. She then formed it by keeping the inner surface of one mandible in contact with the edge of the envelope while the other mandible was repeatedly closed; along with subtle head movements this action apparently pushed resin against the concave inner surface of the other mandible (Figure 1D). Although sticky to human hands, the resin did not stick to the bee. Within a given session of construction, the bee did not extend the envelope only at a single spot, or extend the envelope in a continuous manner, but moved about the envelope between visits to the resin cache, from one side to the other with short bouts of resin-shaping (Figure 2). When females added new material to the previous work there was a thin band of slightly thicker resin that was visible externally as a ridge, which was then extended to a thinner layer by squeezing the resin with the mandibles (a video clip of nest construction behavior is given in the Supplemental Materials). Early on the 4th day after the nest was discovered the envelope was nearly completed, including a small roof defining the top of the entrance hole (Figure 1C); we infer that nest envelope construction takes approximately 6 days. Intermittent observations show that cell construction is completed in less than 2 (1 cell) or 3 days (4 cells).
Much of the very limited information available on orchid bee nest construction comes from studies of cavity nesters in artificial observation boxes, so few comparisons are possible. Euglossa hyacinthina offers an excellent opportunity for detailed studies of building behavior in corbiculate bees with relatively simple architecture, for comparisons with Apis, for which architecture reaches its zenith in the perfect geometric structure of honey bee comb (

A Interior view of a nest of Euglossa hyacinthina under construction. The dark area in the center is where the nest envelope is attached to the plant stem. The numerous droplets are caches of resin (white arrow). B The same nest the next day. C The same nest nearly completed, 3 working days after the photo in A. The white arrow points to the eave over what will become the entrance hole. D A female orchid bee shaping resin along the rim of the growing nest envelope.
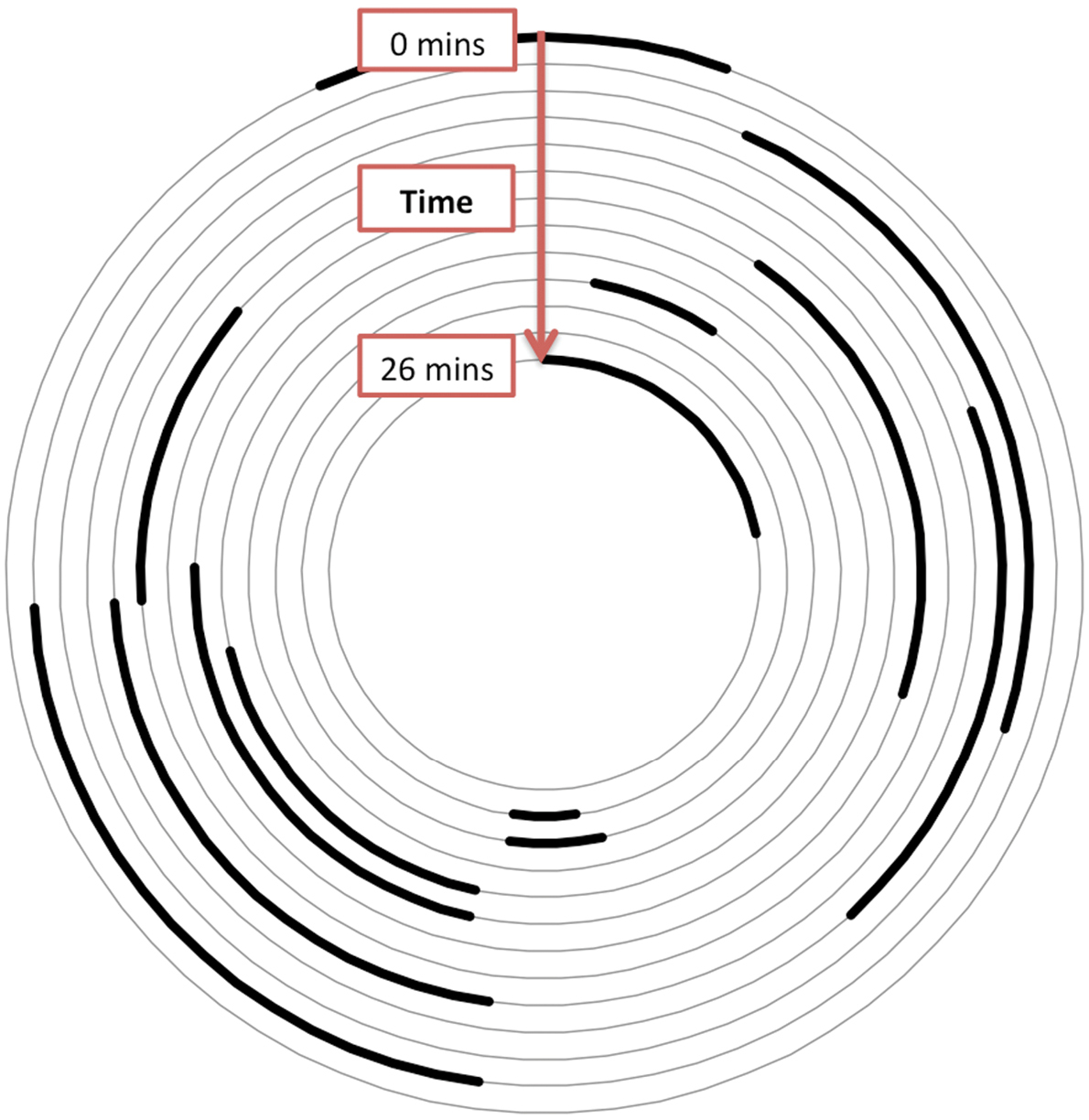
Graphical representation of work effort showing how the bee moves from one side of the nest to another while constructing the envelope. Each arc represents the area the bee worked in a given work session, each lasting approximately from 1 to 2 min. The concentric arcs represent 13 sequential work bouts during a 26 min session of construction; the outer-most arc is the starting position at the beginning of the session and the inner-most one is the ending position.
We thank the Smithsonian Tropical Research Institute (STRI) support staff for logistic help, Donna Conlon for the loan of camera equipment, and the Autoridad Nacional del Medioambiente of Panama for research and collecting permits. The research was supported by general STRI funds to WTW. KEI was supported by a grant from the Arizona State University-Smithsonian Institution collaboration.
A Quicktime video of a female Euglossa hyacinthina working on the construction of her nest envelope. doi: 10.3897/JHR.29.4067.app