





(C) 2013 John T. Huber. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new genus and species of fairyfly, Tinkerbella nana (Hymenoptera: Mymaridae) gen. n. and sp. n., is described from Costa Rica. It is compared with the related genus Kikiki Huber and Beardsley from the Hawaiian Islands, Costa Rica and Trinidad. A specimen of Kikiki huna Huber measured 158 μm long, thus holding the record for the smallest winged insect. The smallest size possible, as measured by body length, for flying insects and wingless arthropods is discussed.
Mymaridae, Tinkerbella, Kikiki, Dicopomorpha, Alaptus, smallest insect, smallest arthropods
The family Mymaridae (Hymenoptera) includes the smallest known insects.
Here we describea new genus of Mymaridae from Costa Rica with specimens below 250 μm in length. We compare it with specimens of Kikiki, a genus described first from the Hawaiian Islands (
Specimens were collected in Costa Rica by JSN using a heavy triangular sweep net fitted with a galvanized metal screen with 4 mm mesh over the opening. Sweeping was done by gently dragging the net through vegetation and dumping the net contents at five minute intervals into 80% ethanol in a sturdy polythene bag to reduce damage to insects in the debris. Sweeping was done for two hours periods (barring rain) so different samples could be compared, if necessary. The samples were sorted later in a laboratory using the method described here to ensure that all the smallest Hymenoptera were found. The procedure is:
1 Stir the entire sample in 80% ethanol very gently to cause the smaller insects to float up from the bottom.
2 Using a 3 ml teaspoon, take enough of the floating material to fill it (a slight stir allows one to gently place the teaspoon into the sample to allow material to settle on to it).
3 Add ethanol from the sample jar to a sorting dish to a depth of 2–3 mm. The sorting dish used in this case consisted of a 9 cm plastic Petri dish with grooves scored at 1 cm intervals on the outside and made visible by drawing black lines in India ink.
4 Examine the sorting dish contents under a binocular microscope by gently moving material across with a pair of forceps and extracting the insects desired. When finished, pour the sorted waste into a 500 g jar.
5 Repeat steps 1–4.
6 After a few repetitions gently stir the jar containing the waste ethanol, allow it to settle for 10 seconds, then pour the supernatant gently into the sorting dish as in step 3 and scan for any remaining small insects.
7 When the unsorted sample has too little ethanol to stir it properly, gently stir the waste jar, allow it to settle for 10 seconds, and use the supernatant ethanol to top up the unsorted sample.
8 If there is not enough ethanol for step 8, add fresh ethanol to the waste jar, stir it slightly, and repeat step 6.
Specimens retrieved were critical point dried and card mounted. Photographs were taken with a digital scanning camera attached to a microscope, and the resulting layers combined electronically using Syncroscopy Auto-Montage™ and, except for primary types, retouched as needed with Adobe™ Photoshop. Micrographs were taken with a Philips environmental scanning electron microscope (ESEM) from uncoated specimens on their cards. A few specimens were slide mounted, either with prior clearing in 10% KOH or without clearing.
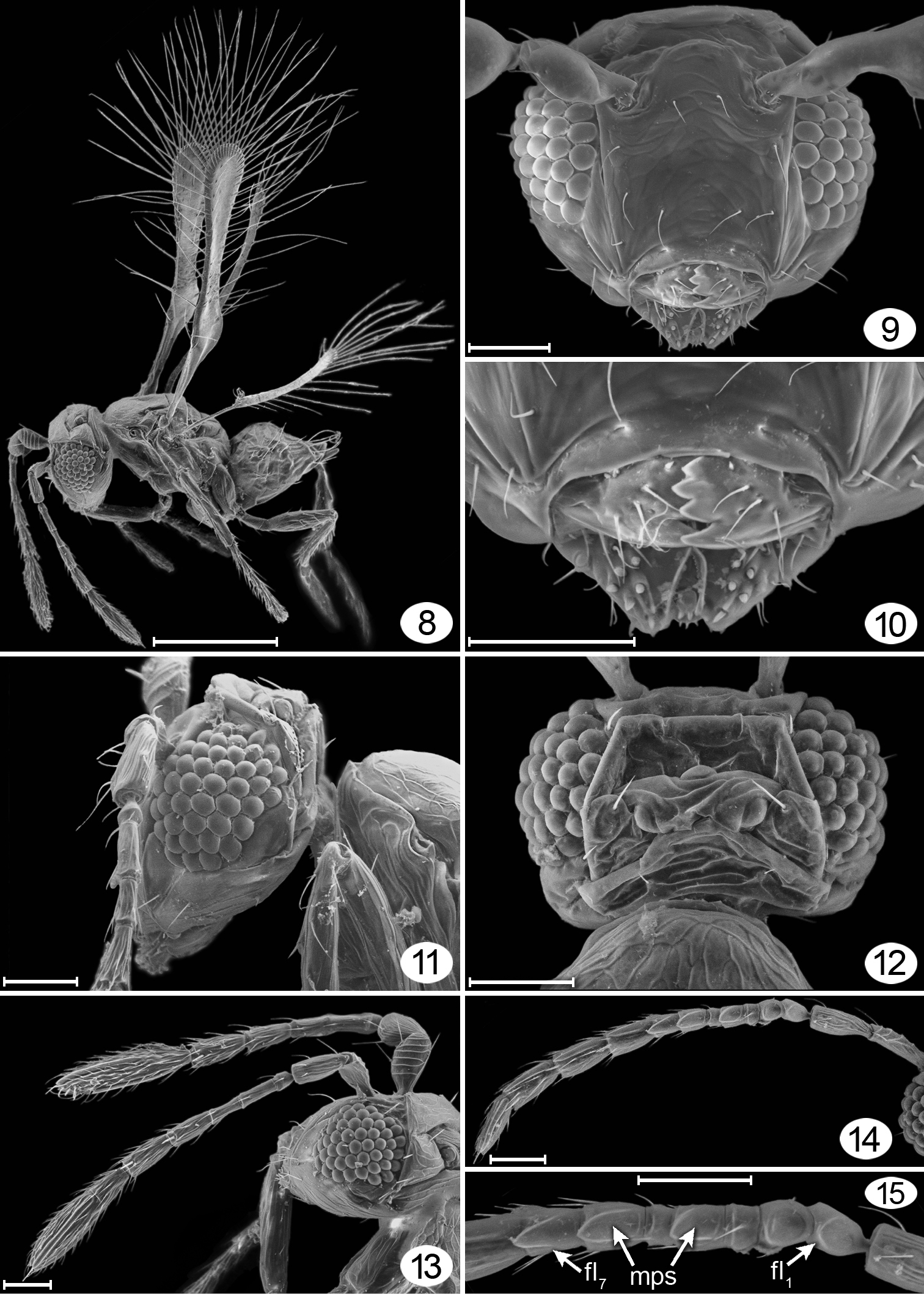
Morphological terms are according to
BMNH Department of Entomology, The Natural History Museum, London, UK.
CNC Canadian National Collection of Insects, AAFC, Ottawa, ON, Canada
INBio Instituto National de Biodiversidad, San José, Costa Rica.
UCRC University of California, Riverside, CA, USA.
urn:lsid:zoobank.org:act:E3F95FC3-C247-41D3-90F9-47C7724EA7E3
Tinkerbella nana Huber and Noyes.
After the fairy Tinker Bell in the 1904 play “Peter Pan” by J.M. Barrie. Gender: feminine.
Tinkerbella is defined by the following combination of features: body length at most about 250 μm; female antenna with funicle 5-segmented and clava entire (Figs 1, 3, 5, 13), compound eye with about 50 ommatidia (Figs 11, 13), and tarsi 4-segmented (Fig. 22).
Kikiki is the most closely related genus to Tinkerbella. It is distinguished from Tinkerbella by the following combination of features: female with funicle 4-segmented and clava 2-segmented (Figs 24, 37, 38), compound eye with about 20–25 ommatidia (Figs 24, 38, 39, 40), and tarsi 3-segmented (Fig. 47, 48).
Female. Body minute, at most about 250 μm long. Head in dorsal view (Fig. 12) about 1.6× as wide as long, in lateral view (Fig. 11) about 1.6× as high as long, and in anterior view (Figs 5, 9) about 1.3× as wide as high. Face slightly longer than wide and ventrally separated from clypeus by curved epistomal sulcus. Toruli about their own diameter from transverse trabecula. Clypeus apparently extending entire width of mouth opening, with anterior tentorial pits visible, slit-like (Fig. 10). Mandible with 4 teeth, the lower one more distinct than the upper three (Fig. 10). Eye with about 50 facets, in lateral view about 1.4× as high as long and not extending to back of head dorsally. Malar distance about 0.4× eye height, with straight malar sulcus. Vertex at about right angle to face above toruli, with supraorbital trabecula divided medially into two parts. Ocelli in normal triangle with POL: LOL: OOL about 1.1: 0.5: 0.5, enclosed by conspicuous rectangular stemmaticum and with sulci extending laterally to endpoints of posterior section of supraorbital trabecula (Fig. 12); occiput divided by transverse sulcus just dorsal to foramen medially and ventral to eyes laterally (Fig. 17). Antenna. Scape about 3.9× as long as wide, with faint longitudinal reticulations on outer surface and distinct transverse ridges on inner surface (Figs 5, 13). Pedicel about 0.2× as long as scape. Funicle 5-segmented. clava entire (a faint, partial division visible, however) (Figs 5, 13). Mesosoma. About 1.34× as long as high and about 1.3× as long as wide. Pronotum thin, in dorsal view scarcely visible (Fig. 7), in lateral view with large lateral panel extending posteriorly to level of wing base (Fig. 11). Mesoscutum (Figs 16, 17) slightly longer than scutellum, with deep notauli. Anterior scutellum and axilla completely separated by deep straight transverse sulcus from slightly longer, longitudinally divided frenum. Second phragma projecting to apex of propodeum. Metanotum with dorsellum distinct, about 0.5× as long as frenum, apparently divided by a median longitudinal groove (Figs 16, 17), and much wider than very narrow lateral panel of metanotum (Figs 16, 17). Mesopleuron (Fig. 17) divided by fairly straight, faint transepimeral sulcus into narrower mesepisternum and wider mesepimeron. Metapleuron triangular. Propodeum in dorsal view (Fig. 16) with anterodorsal area mostly smooth, posterodorsal area with minute rounded spicules medially, with propodeal seta about midway between spiracle and posterior margin of propodeum; in lateral view (Fig. 18) strongly sloping. Propodeal spiracle circular, smaller than pronotal spiracle and distinctly separated from anterior margin of propodeum. Wings. Fore wing (Figs 6, 8, 20) narrow, almost parallel sided, with posterior margin distinctly concave beyond pronounced lobe (Fig. 21) at level of parastigma; wing surface bare except for fewer than about 10 scattered microtrichia on dorsal and ventral surfaces; fringe setae much longer than wing width. Venation extending almost 0.7× fore wing length; submarginal vein about 1.6× parastigma length, and parastigma about 0.5× marginal + stigmal vein lengths (their relative proportions about 24/15/29); hypochaeta close to proximal macrochaeta (Fig. 21); parastigma with distal macrochaeta about 3× as long as proximal macrochaeta (Fig. 21). Hind wing (Figs 6, 8) curved, with anterior margin concave, paralleling convex posterior margin, apex bluntly rounded, with long fringe setae at apex and along posterior margin to just beyond level of relatively large hamuli; wing surface bare except for row of fewer than about 8 microtrichia along anterior margin except toward hamuli where they are located more posteriorly. Legs. Tarsi 4-segmented, tarsomere 1 slightly shorter than remaining tarsomeres, and tarsomere 4 distinct, as long as tarsomere 3 (Fig. 22). Metasoma. Slightly shorter than mesosoma (critical point dried specimens) (Fig. 7); petiole very short (scarcely visible), about 4.3× as wide as long, but clearly narrower than gt1; gaster in dorsal view distinctly narrower anteriorly than medially (Fig. 7) with gt6 the longest tergite and apparently without a spiracle. Ovipositor (Figs 7, 18) as long as gaster, slightly exserted beyond gastral apex. Cerci with 3 long setae extending just beyond ovipositor apex (Fig. 19).
Male. Antenna 11-segmented (Fig. 14), with 2 mps each, except fl3 and fl5 much shorter and without mps (Fig. 15), the mps decreasing in width on each segment, from almost circular on fl1 to linear on fl11.
urn:lsid:zoobank.org:act:EFCB279C-935F-4098-873F-4B7C5C935E8F
http://species-id.net/wiki/Tinkerbella_nana
Figs 1, 3 – 22♀ (INBio) on slide labelled, 1. “COSTA RICA: Heredia, La Selva, 75m, 10°26'N, 84°01'W, 27–28.ii.2003, J.S. Noyes, sweeping, cleared in clove oil, mounted in Canada balsam”. 2. “Tinkerbella nana Huber and Noyes ♀dorsal Holotype”.
Paratypes. 7♀ and 2♂. COSTA RICA: Alajuela. Reserva Rincón Forestal, Estación Caribe, 400m, 10°53'N, 85°18'W, 400m, 19–22.ii.2003, J.S. Noyes (1♀, 1♂, CNC); Arenal National Park, sendero Pilón 10°27'N, 84°43'W, 600m, 26.ii.2003, J.S. Noyes (1♀, CNC). Heredia. Same data as holotype (5♀, 1♂, CNC, BMNH, InBio).
After the dog Nana in Peter Pan and coincidentally from nanos, the Greek word for dwarf. Treated as a (feminized) noun in apposition.
Female. Body length 225–250 μm (n=6). Colour. Very pale, the scape, pedicel, pronotum laterally, gaster laterally, and legs except apical tarsomere sometimes lighter, almost white; head, mandibles, mesoscutum, anterior scutellum and propodeum with a pale yellow or pale brown tinge, occasionally head and mesosoma, especially mesoscutum, more uniformly and extensively brown; trabeculae and a minute spot next to fore wing base dark brown; eyes and ocelli distinctly reddish (Fig. 3). Fore wing with brown tinge behind most of venation except its apex (Fig. 6). Hind wing fairly uniformly light brown from just before hamuli to slightly lighter apex. Head. 65–99 wide, with transverse reticulate sculpture on face (Fig. 9), vertex (Fig. 12), and occiput (Fig. 12). Antenna. Flagellum (Figs 5, 13) with 1 mps on fl4, 1 mps on fl5, and 4 mps on clava. Fl1–clava length/width (n=2–width or 4–length): scape, 25–30/13–14, pedicel 19–27/17–18, fl1 8–10/7–8, fl2 12–21/6–7, fl3 11–16/7–8, fl4 8–24/9, fl5 23–31/10–11, clava 59–65/15–17. Mesosoma. Mesoscutum (Figs 7, 16, 17) with slightly longitudinal, reticulate sculpture and raised meshes. Scutellum smooth anteriorly, frenum slightly wrinkled. Dorsellum smooth to slightly wrinkled. Wings. Fore wing with a few scattered microtrichae behind venation from proximal macrochaeta on, and 3 or 4 just beyond venation (Fig. 6). Fore wing length 195–240 (n=5, card mounts at 200×), width 30–50, longest marginal setae 100–155; hind wing length 145–200, width 5–10, longest marginal setae 105–130. Metasoma. Petiole 34 wide, 7 long (n=1). Gaster with segments somewhat wrinkled and sometimes transversely creased posteriorly (Figs 7, 18); gt2–gt6 each with a few fairly long, suberect setae dorsally and laterally.
Male. Body length 210–230 μm (n=2). Colour light brown (Fig. 4). Fl3 and fl5 the shortest segments, less than half length of the segment following and without mps (Fig. 15).
Unknown. The specimens from Alajuela were collected by sweeping in fairly young secondary forest (20 years old maximum) mixed with a small amount (ca. 1 ha) of primary forest.
Costa Rica.

Habitus, lateral. 1 Tinkerbella nana female 2 Kikiki huna, female. Scale line = 100 μm.

Tinkerbella nana female. 3 holotype, uncleared on slide 4 paratype, critical point dried on card. Scale line = 100 μm.
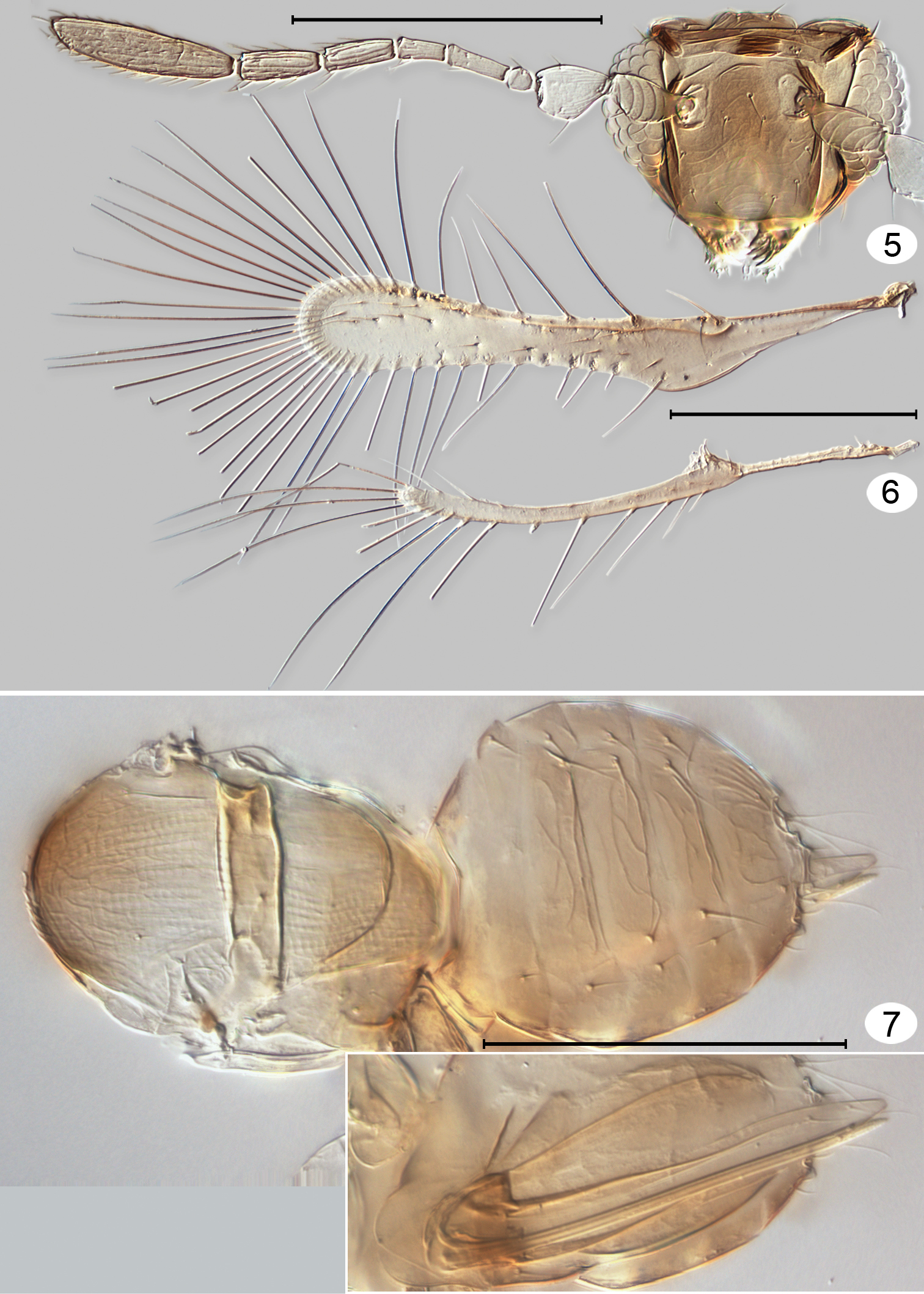
Tinkerbella nana female paratype, cleared on slide. 5 head + right antenna, anterior 6 wings 7 mesosoma + metasoma dorsal, and ovipositor ventral (inset). Scale line = 100 μm.
Tinkerbella nana paratype female (except 14, 15), micrographs. 8 habitus, lateral 9 head, anterior 10 mouthparts, anterior 11 head + pronotum, lateral 12 head, dorsal 13 head + antennae, lateral 14 male antenna, dorsal 15 male antenna, pedicel–fl8, dorsal. Scale line = 20 μm, except Fig. 8 = 100 μm.
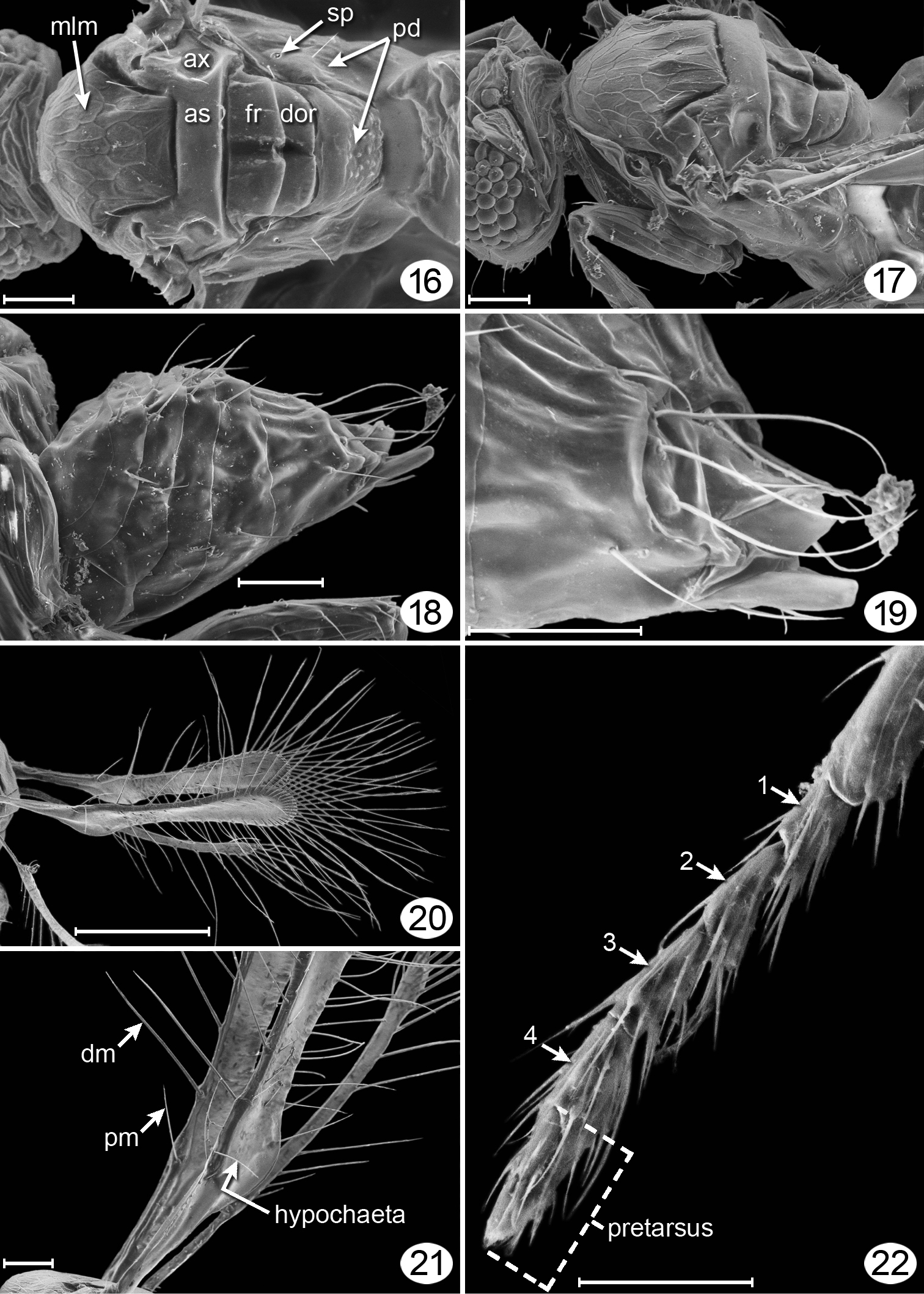
Tinkerbella nana female paratype, micrographs. 16 mesosoma, dorsal 17 mesosoma + back of head, dorsolateral 18 metasoma, lateral 19 apex of gaster, dorsolateral 20 wings (left wing ventral surface, right wing dorsal surface) 21 wing, basal half 22 left mesotarsus. See appendix for abbreviations. Scale line = 20 μm, except Fig. 20 = 100 μm.
http://species-id.net/wiki/Kikiki
Specimens of Kikiki from Costa Rica were collected and examined since the original generic description based on eight specimens from the Hawaiian Islands (
Female. Head.Face about 1.7× as high as wide, slightly depressed medially (Fig. 24), separated ventrally from oral cavity by distinct epistomal suture. Clypeus transverse, in same plane as face, narrowly oval and extending entire width of oral cavity, with anterior tentorial pits visible sublaterally (Fig. 37). Mandible with lower tooth separated from remaining teeth by deeper notch than notches separating the teeth above it. Vertex (Figs 30, 40) laterally with well defined supraorbital trabecula divided medially; ocelli enclosed by a distinct, rectangular stemmaticum, the vertex thus divided into anterior (smooth) and posterior (faintly sculptured) areas by a transverse groove extending between eyes from just anterior to each supraorbital trabecula and along anterior margin of slightly triangular mid ocellus; lateral ocelli almost vertical, facing away from each other. Occiput separated from vertex by slightly curved groove extending between posterior apex of each supraorbital trabecula and almost touching lateral ocelli (Figs 40, 41), and occiput divided into dorsal and ventral areas by curved groove extending between lower margin of eyes and above foramen (Figs 25, 42). Head, except vertex anteriorly, with faint, reticulate sculpture. Mesosoma. Pronotum with a slightly crenulated dorsal margin (Figs 40, 42). Mesoscutum (Figs 26, 42, 44) with notauli narrow near anterior apex, distinctly widening (narrowly triangular) posteriorly to the medially deep and wide transscutal articulation. Scutellum with axilla strongly advanced anteriorly into lateral lobe of mesoscutum. Frenum apparently divided mediolongitudinally by a wide depression (Figs 44, 45). Mesosoma except anterior scutellum with faint, reticulate sculpture. Legs. Fore leg with bifurcate strigil and calcar with 4 rounded teeth, separated from one another by almost their own diameter (Fig. 34-male). Metasoma. Petiole (Fig. 26) extremely short and distinctly narrower that gt1. Terga each with a median transverse fold extending across tergum medially or nearer posterior margin (Figs 28, 45, 46).
Male. Body 235 μm from transverse trabecula to gastral apex (Fig. 30). Antenna with 9segmented funicle (Fig. 33;
The male is described from a single slide-mounted specimen from Australia. No males have yet been collected in the New World. Whether the larger body and eyes of the Australian specimen are sex differences or indicative of a species different from Kikiki huna cannot be determined until females are obtained from the same locality in Western Australia and compared with the Hawaiian and Neotropical specimens.
Female. Body length (critical point dried specimens) 158–190 μm (n=10). Antenna. Funicle segments and basal claval segment without mps, apical claval segment with 3 mps (Figs 24, 43). Antennal length/width measurements (n=4, Costa Rica specimens): scape, 36–47/10–11, pedicel 19–25/12–13, fl1 8–9/5–6, fl2 16–18/6, fl3 14–17/6–7, fl4 12–18/7–8, clava 46–54/15. Wings. Fore wing (Fig. 27) length 182–226, width 20–24, length/width 9.10–9.24, longest marginal setae 102–123 (n=3, slide mounts), hind wing (Figs 23, 27) length 162–198, width 4–5, longest marginal setae 96 (n=1–3).
Male. Unknown for Neotropical region.
20♀. COSTA RICA. Heredia. La Selva Biological Station, 10°26'N, 84°01'W, 75m, 27–28.ii.2003, J.S. Noyes (1♀, CNC). Puntarenas. La Gamba Biological Station, 8°42'N, 83°12'W, 150m, 13–14.ii.2006, J.S. Noyes (1♀, BMNH); Reserva Absoluta Cabo Blanco, 9°35'N, 85°36'W, 30m, 16–17.ii.2009, J.S. Noyes, sweeping (9♀, CNC, INBio, UCRC); Reserva Privada Karen Morgensen, 9°52'N, 85°03'W, 305m, 23–24.ii.2007, J.S. Noyes, sweeping (6♀, BMNH). HAWAIIAN ISLANDS. Molokai I.: Mapulehu (1♀ paratype, CNC). TRINIDAD & TOBAGO. Trinidad. Curepe, Santa Margarita Circular Road, 8.xii.1974–2.ii.1975, F.D. Bennett (2♀, CNC).
We cannot find any morphological differences suggesting that the specimens from Costa Rica and Trinidad are different from the Hawaiian specimens. The number and distribution of mps on the antennal segments as reported by
At the genus level, and even the species level, the Hawaiian fauna at low elevation appears to be almost entirely represented by exotic species (
Tinkerbella would key to Kikiki in
| 1 | Venation long, clearly more than half wing length; tarsi 3- or 4-segmented | 1a |
| – | Venation short, clearly less than half wing length; tarsi 5-segmented | couplet 2 in |
| 1a | Clava 2-segmented, funicle 4-segmented; tarsi 3-segmented | Kikiki Huber and Beardsley |
| – | Clava entire; funicle 5-segmented, tarsi 4-segmented | Tinkerbella Huber and Noyes |
However, several features of both genera show that their placement in the Alaptus group of genera is wrong and that they are best placed in the Anagrus-group because they share at least seven features with some or all of those genera: frenum apparently longitudinally divided by a groove, petiole and base of gaster distinctly narrower than propodeum and middle of gaster, tarsi with at most 4 tarsomeres (3 in Kikiki), stemmaticum present, mandibles with 4 teeth, fore wing venation with proximal macrochaeta much shorter than distal macrochaeta, and second phragma not projecting past posterior margin of propodeum.
Both Kikiki and Tinkerbella are distinguished from other Anagrus-group genera by the venation clearly longer than half the wing length and hind wing essentially without fringe setae on the anterior margin. The other genera in the group have the venation clearly less than half wing length and the hind wing fringed with fairly long setae on the anterior margin. No member of the Anagrus group of genera is anywhere near as small as Tinkerbella or Kikiki so these genera can be distinguished on body size alone.
Some members of the Alaptus or Camptoptera groups of genera are also extremely small but except for Alaptus borinquensis none has been found that equal minute size of Tinkerbella or Kikiki, i.e., less that about 250 μm. One specimen of Alaptus from the same collecting event in Costa Rica as Kikiki huna measured 272 μm and species of Eofoersteria (also with 4-segmented tarsi, despite being in the Camptoptera group) are about 320 μm. The six specimens (4 females, 2 males, on 4 slides, USNM) of Alaptus borinquensis are uncleared and mounted mostly in lateral view in Canada balsam, and some are slightly shrivelled. The smallest specimen, a male, measured 203 μm in length, not 186 μm as reported by
What is the smallest size that an adult insect can attain, as measured by body length, and/or fore wing length for flying insects? If something is physically possible in living things some individuals of at least one species, extinct or extant, will likely have achieved it. So the lower size limit, by whatever measure of size is chosen, was almost certainly already evolved—somewhere, sometime. If we have not already found them, we must surely be close to discovering the smallest insects and other arthropods.
The minimum size possible for invertebrates with articulated appendages of locomotion (superphylum Arthropoda) that would allow crawling, walking, or active flight (wing flapping, not gliding) is determined by two types of constraint: internal (physiological and structural) and external (physical). For multicellular animals, the size and structure of cells cannot go below a certain minimum before they cannot function for the purpose intended. Therefore, once any type of cell has attained its minimum size, the number of cells making up a particular tissue or organ must decrease as the animal becomes smaller. But there evidently is a lower limit to cell number in any particular organ for it to function as intended, perhaps as low as one, e.g., a single muscle fiber or ommatidium. Or the tissue or organ may be dispensed with entirely, usually because it is no longer necessary, e.g., eye loss in obligate cave-dwelling species (troglobites). Once these internal constraints are reached the arthropod cannot become any smaller.
As body size decreases, external constraints, e.g., desiccation (
Even with the increased mechanical efficiency resulting from smaller body size and energy conservation efficiencies conferred by Weis-Fogh clap-and-fling flapping (
A diversity of other very small insects capable of active flight have the wing surface (membrane) reduced and wing fringes, especially of the hind wing, greatly lengthened, e.g., species of Thysanoptera, various families of small Lepidoptera and parasitic Hymenoptera, and Ptiliidae (Coleoptera). While this may slightly reduce wing weight the reduced wing surface and relatively long setae are more likely to have an aerodynamic function, perhaps to reduce turbulence and hence drag on a wing flapping at several hundred beats per second. Interestingly, in Kikiki (Fig. 27), Tinkerbella (Fig. 6) and Megaphragma (Fig. 51) the fringe setae along the leading edge of the hind wing are absent or almost so, whereas in other genera of Trichogrammatidae and Mymaridae they are present, albeit short. The separation of muscles that power flapping, i.e., the indirect flight muscles that cause thoracic distortion, resulting in wing flapping, from the direct flight muscles that control wing pitch may also be important in allowing insects capable of active flight to attain a minute body size.
Relatively long legs powered by sufficiently strong muscles to elevate the body may be linked to and necessary for active flight. The muscles required to move articulated legs to enable walking by lifting the entire body off the ground and moving forward must have a lower size limit or the legs could not be used for that purpose. Two sets of opposing intrinsic muscles are needed: extensor/elevator muscles strong enough to lift the entire body sufficiently above the substrate to initiate flight, perhaps by allowing a wing stroke of more than 90°, and flexor/depressor muscles to allow walking, and perhaps secondarily to draw the legs against the body during flight to reduce drag. High speed photography of Trichogramma lifting off a substrate (
In insects with free living adults and larvae the lower body length limit seems to be about 400 μm (
The constraint of minimum egg size as a determinant of minimum body size does not apply to parasitic insects. Eggs in these can be much smaller because the larvae hatch inside the host. They do not have to search actively for food because it completely envelops them. The limiting factor to small size in adult parasitic insects must therefore be minimum cell size and sufficient cells of each type of tissue, as discussed above. Females of Tinkerbella, Kikiki, and Alaptus borinquensis are the smallest Mymaridae, and some specimens of Kikiki are the smallest recorded winged insects. The body length of five specimens from Costa Rica is 158 μm (1 female), 160 μm (2 females), and 170 μm (2 females). Three specimens are thus smaller than the smallest recorded females of Megaphragma caribea, the previous record holder at 170 μm (
The next step in body length reduction in insects is in Dicopomorpha echmepterygis. Females are winged and relatively long (one measured 386 μm dry but not shivelled and 550 μm on a slide mount), but the males slide-mounted males were 139–240 μm long (n=8) (
Further reductions in body length occur in terrestrial Arthropoda other than insects. In mites, the smallest adult individuals of several species in three families are less than 95 μm in length: Cochlodispus minimus Mahunka at 79 μm (
For comparison with terrestrial arthropods, larvae of the marine parasites of Copepoda Stygotantulus stocki Boxhall and Huys at 94 μm (
Below a certain body length it is useless to have articulated appendages because the segments could not be moved relative to one another, or the entire appendage relative to the body, by intrinsic muscle power alone. Instead, if appendages of locomotion exist at all (e.g., pseudopods), they would be short and wide, would not be articulated, and would be moved instead by body muscles causing hydrostatic changes in pressure, combined perhaps with flexor muscles originating within the body but attached near or at the appendage apex. The length of the larva of Neoliochthonius piluliferus rivals some Rotifera, also as short as 50 μm, suggesting that at about this size the changeover from locomotion by partial muscle power intrinsic to leg segments (Acari) to hydrostatic power alone (Rotifera, other non-Arthropoda) may occur.
We suggest that the smallest winged insects capable of flapping flight could not be less than about150 μm in length, and the smallest capable of normal walking (body lifted entirely of substrate) not below about 125 μm. Among insects, Kikiki huna may well have attained the lower limit for active flight and Dicopomorpha echmepterygis the lower limit for normal walking. Among other arthropods capable of walking, 80 μm is suggested as the lower limit for adults and ca. 50 μm for immatures.
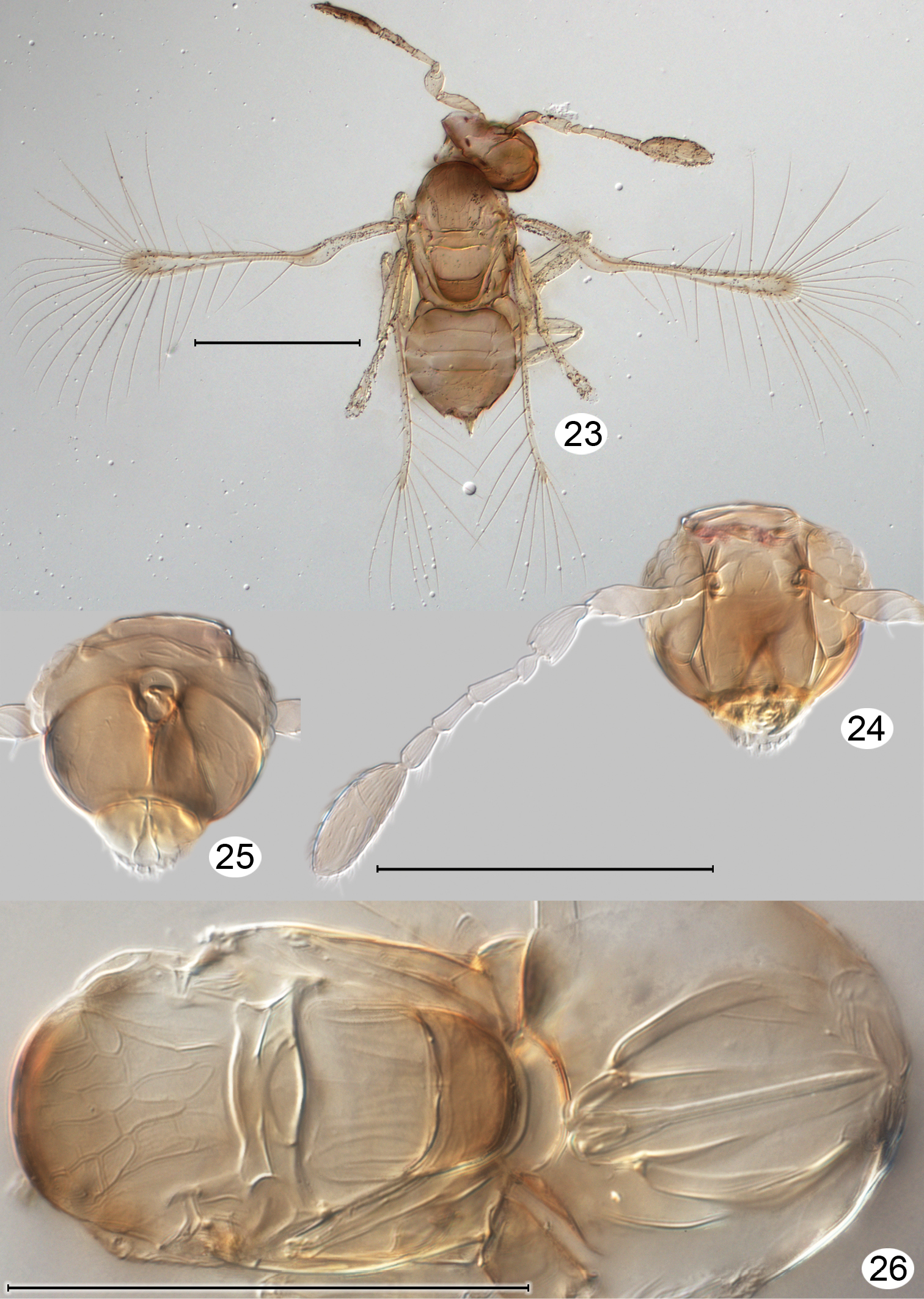
Kikiki huna female, on slide (cleared, except Fig. 23). 23 habitus, dorsal 24 head + right antenna, anterior 25 head, posterior 26 mesosoma, dorsal + metasoma, dorsal but focus at lower plane to show ovipositor. Scale line = 100 μm.

Kikiki huna female, cleared on slide. 27 wings 28 metasoma, dorsal surface 29 metasoma, ventral surface (seen dorsally through cleared metasoma). Scale line = 100 μm.
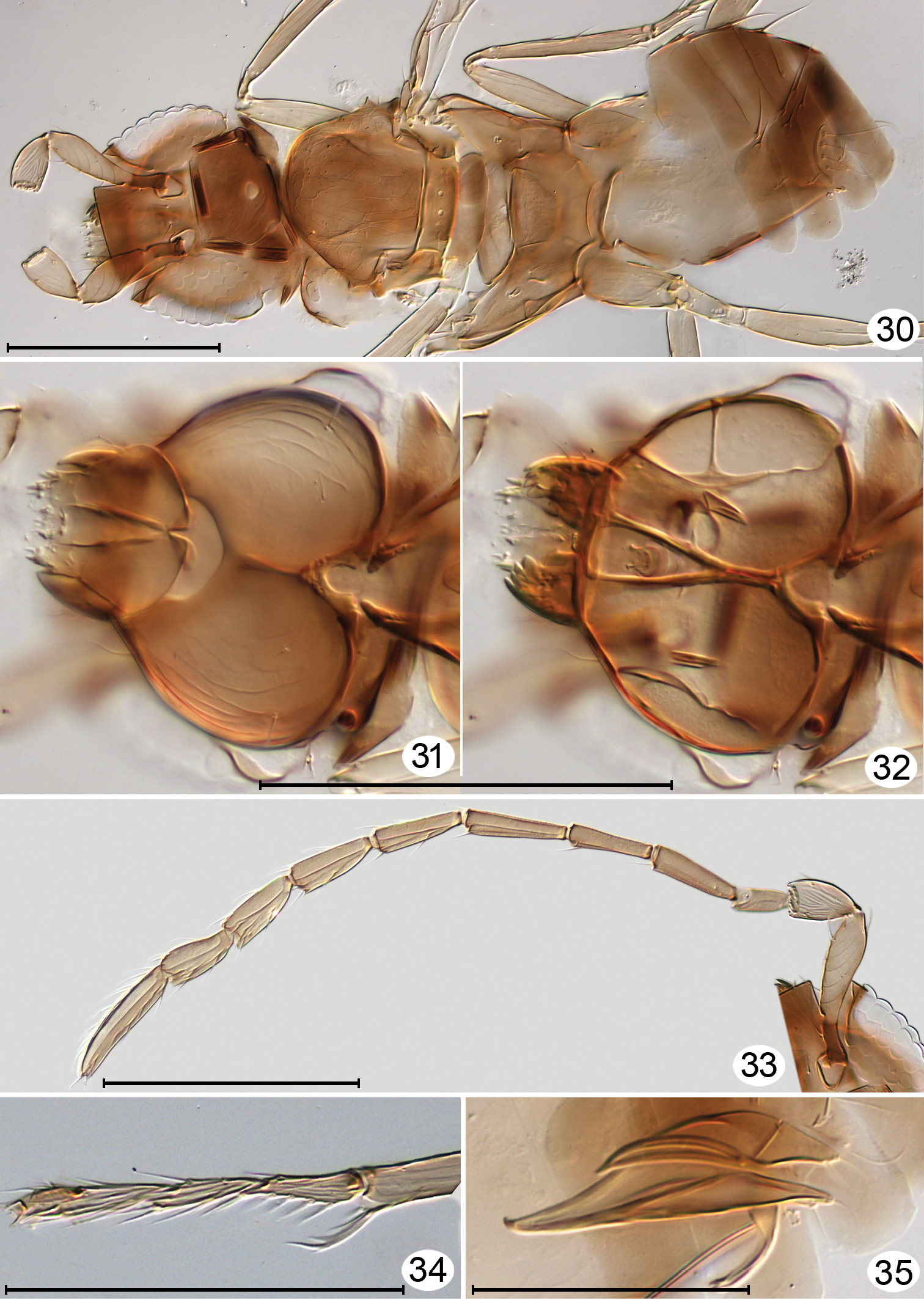
Kikiki sp. male, cleared on slide. 30 head anterodorsal + mesosoma and metasoma, dorsal 31 head, posterior 32 head from anterior, focused at different plane to show internal skeleton and mandibles 33 antenna 34 protarsus 35 genitalia, dorsolateral. Scale line = 100 μm, except Figures 34 and 35 = 50 μm.

Kikiki huna female, micrographs. 36 habitus, dorsal 37, head + antennae, anterior 38 habitus, lateral 39 head, lateral 40 head + anterior mesosoma, dorsolateral 41 head + base of antenna, dorsal 42 head, posterodorsal + anterior mesosoma, dorsal. Scale line = 20 μm, except 36, 38 = 100 μm.
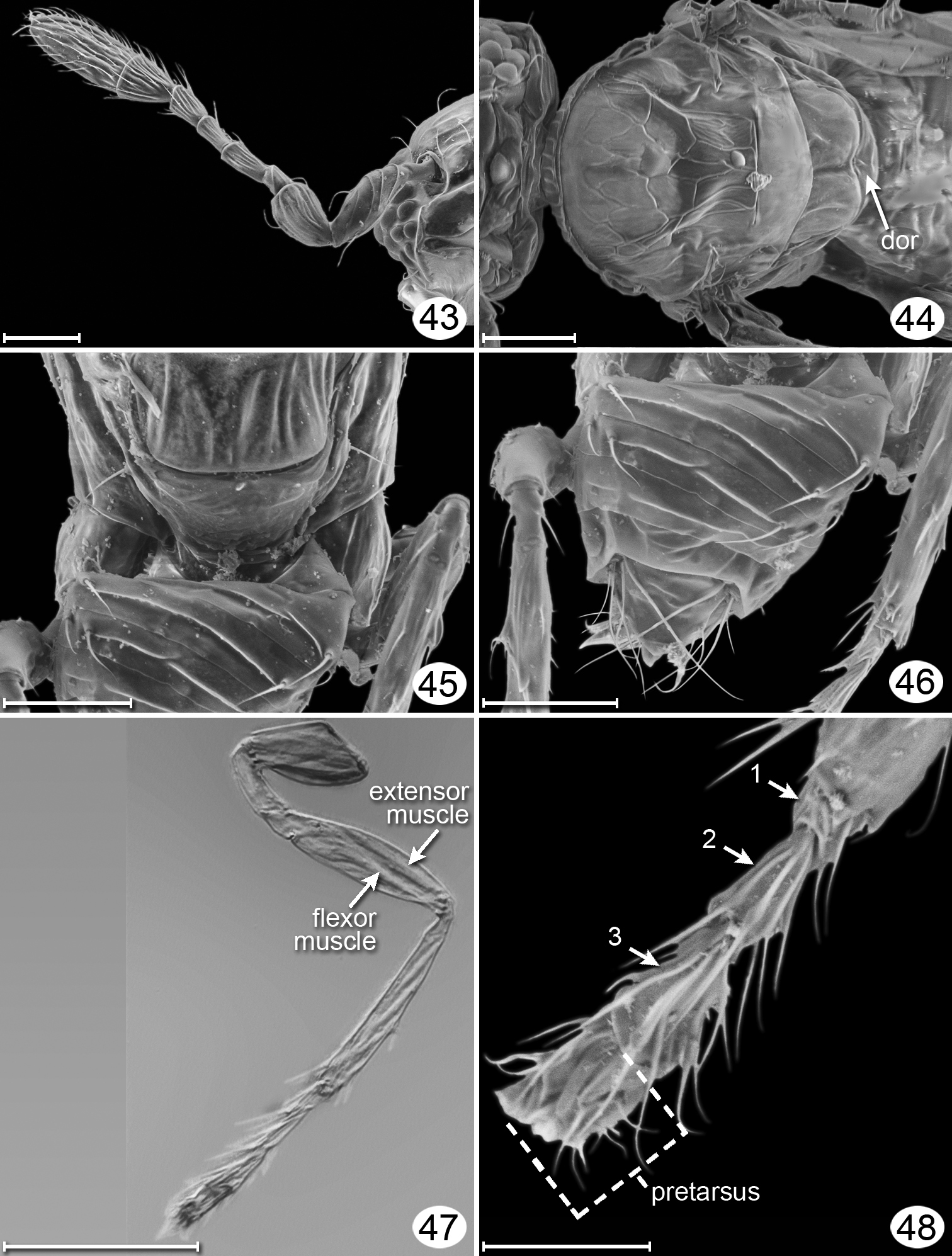
Kikiki huna, female, micrographs except Fig. 47. 43 antenna, lateral 44 mesosoma, dorsal 45 frenum – anterior half of metasoma, dorsal 46 metasoma, dorsal 47 hind leg, uncleared lateral, showing muscles; 48, right metatarsus, dorsal. Scale line = 20 μm, except Fig. 47 = 50 μm and 48 = 10 μm.

Megaphragma sp., uncleared on slide. 49 head + antenna, anterior (note black eyes) 50 mesosoma + metasoma, dorsal (note huge mesophragma, hence the genus name) 51 wings + middle leg (note 3-segmented tarsi, diagnostic for Trichogrammatidae). Scale line = 100 μm.
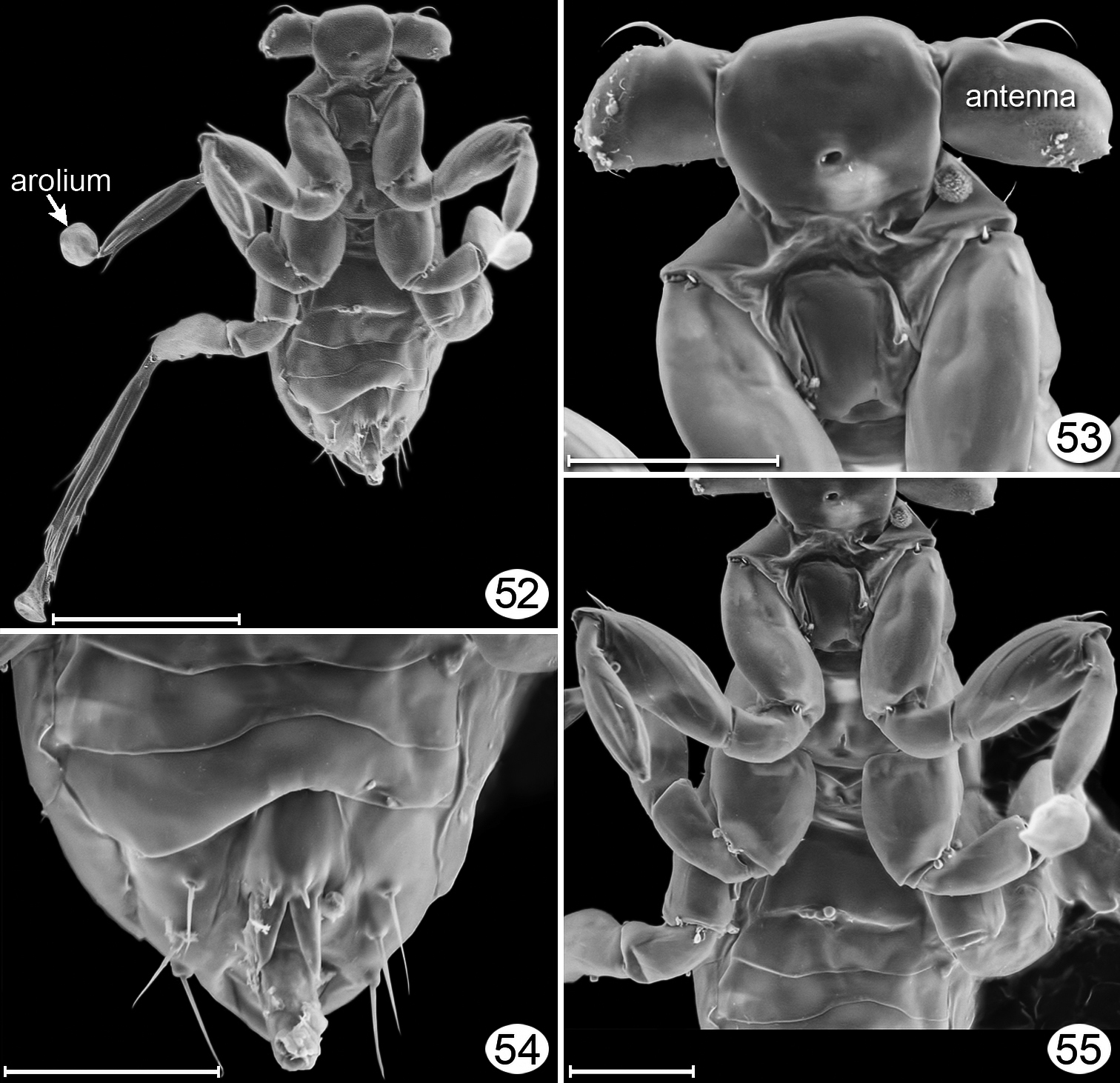
Dicopomorpha echmepterygis male, paratype, ventral. 52 habitus 53 head + prothorax + procoxa 54 apex of gaster 55 mesosoma + base of most legs and metasoma. Scale line = 20 μm, except Fig. 52 = 50 μm.
We thank J. Read (Natural Resources Canada, Ottawa) for preparing the photographs and micrographs and compiling the plates, the senior author’s brother, P. Huber (Hanover, New Hampshire), for discussions on the effect of air viscosity for very small flying insects, E. Lindquist (Agriculture and Agri-Food Canada, Ottawa) and D. Walter (Royal Alberta Museum, Edmonton) for interesting discussions and information on Acari, and V. Grebennikov (Canadian Food Inspection Agency, Ottawa) for information on publications by A. Polilov. The type series of Alaptus borinquensis was sent to JTH by M. Gates (Systematic Entomology Laboratory, National Museum of Natural History, Washington, DC) and we are grateful for his help.