Research Article |
|
Corresponding author: Simon van Noort ( svannoort@iziko.org.za ) Academic editor: Tamara Spasojevic
© 2023 Terry Reynolds, Simon van Noort.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Reynolds T, van Noort S (2023) Two new species of the genus Cryptopimpla Taschenberg (Hymenoptera, Ichneumonidae, Banchinae) with an updated key to African species. Journal of Hymenoptera Research 96: 667-696. https://doi.org/10.3897/jhr.96.104038
|
Abstract
A revised illustrated key to Afrotropical species of the genus Cryptopimpla Taschenberg is provided, with the inclusion of two new South African species, C. orenji Reynolds & van Noort, sp. nov. and C. hoerikwagga Reynolds & van Noort, sp. nov., which are described and illustrated. The recovery of the first female specimens of Cryptopimpla goci Reynolds & van Noort in samples from Fernkloof and Grootbos nature reserves, and subsequent morphological reassessment of generic affinity based on female characters, no longer supports the placement of this species in Cryptopimpla. The transfer of C. goci to Lissonota Gravenhorst is proposed here: Lissonota goci (Reynolds & van Noort), comb. nov., and the female is described. New Afrotropical distributional records for the previously described Cryptopimpla species are presented and notes on the distribution and diversification of the species are also provided. Online interactive Lucid keys to the 11 Afrotropical Cryptopimpla species are available at: http://www.waspweb.org.
Keywords
Afrotropical region, Atrophini, distribution, Ichneumonoidea, Lucid identification keys, South Africa, species diversity, taxonomy
Introduction
Afrotropical Cryptopimpla Taschenberg, 1863 represent 17% of the world’s Cryptopimpla species (
The hosts of Cryptopimpla species that occur in the Afrotropical region remain unknown. Members of the tribe Atrophini generally attack semi-concealed hosts such as lepidopteran leaf rollers (
Although Cryptopimpla has a worldwide distribution, its species richness in the temperate regions of South Africa (and elsewhere) support relative affinities of the genus to specific biogeographic areas defined by habitat and climate (
In this paper, we update the species key to African Cryptopimpla, reassess the generic affinities of Cryptopimpla goci Reynolds & van Noort, 2016, describe two new species, and provide links to the revised online interactive Lucid pathway and Lucid matrix keys available on WaspWeb at http://www.waspweb.org (
Materials and methods
Photographs
Images were acquired at the
Iziko South African Museum (
Mapping
The distribution maps for the African Cryptopimpla species were produced using SimpleMappr (
Specimen acquisition
Specimens were extracted from bulk inventory survey samples preserved in 96% ethanol and housed in the Iziko South African Museum entomology wet collection that had been sorted to family level. These samples emanate from continuous inventory surveys using a range of collecting methods including Malaise traps, yellow pan traps, yellow funnel traps, pitfall traps, sweeping, Winkler bag extraction of leaf litter and UV light trapping conducted in Africa over the last 31 years by the second author (
Digitization
All specimen data has been digitized into the Iziko South African Museum Specify 6 database.
Depositories
SAMC Iziko South African Museum, Cape Town, South Africa (Curator: Simon vanNoort).
Nomenclature and abbreviations
The morphological terminology follows
B body length, from toruli to metasomal apex (mm);
A antenna length, from base of scape to flagellar apex (mm);
F fore wing length, from tegula to wing apex (mm);
CT (clypeus transversality index) maximum width of clypeus: median height;
ML (malar space length index) malar space (shortest distance between mandible base and compound eye): basal mandibular width;
IO (inter-ocellar index) shortest distance between posterior ocelli: ocellus diameter;
OO (oculo-ocellar index) shortest distance between eye and posterior ocellus: ocellus diameter;
Fln (length index of flagellomere n) length: width of flagellomere n;
OT (ovipositor sheath-tibia index) length of ovipositor sheath: length of hind tibia.
The first three measurements (absolute measures) were measured on all specimens in the type series, with measurements from the primary type reported separately in brackets if necessary.
Identification keys
Lucid pathway and Lucid matrix keys were developed using Lucid Builder version 4.0.37. Character matrices were generated and edited using Microsoft Excel; matrices were then used as input into Lucid matrix key production (
Results
Specimen acquisition and distribution maps
Historically there were two Cryptopimpla specimens in the Iziko South African Museum collection: the holotype of C. rubrithorax Morley, 1916 (collected in 1914) and a specimen of the recently described species C. zwarti Reynolds Berry & van Noort, 2016 (collected in 1990). The remaining 60 specimens were collected by the second author over the last thirty years from many diverse vegetational localities. The majority of the resultant bulk samples have yet to be sorted and we expect that numerous further specimens reside in the unsorted samples, probably at least tripling the number of known specimens. Most of the mobilized specimens were collected in Malaise traps, with a single specimen collected by sweeping and two specimens recovered from yellow pan trap samples. A summary of the abundance and distribution of the species treated here is provided in Table
Number of known specimens, the known provincial distribution and vegetation biome association of African Cryptopimpla species.
| Cryptopimpla species | Known specimens | Recorded distribution | Biome association |
|---|---|---|---|
| C. elongatus | 1 | Northern Cape | Fynbos |
| C. fernkloofensis | 1 | Western Cape | Fynbos |
| C. hantami | 2 | Western Cape | Fynbos |
| C. hoerikwagga | 1 | Western Cape | Fynbos |
| C. kogelbergensis | 6 | Northern & Western Cape | Fynbos; Succulent Karoo |
| C. neili | 1 | Western Cape | Fynbos |
| C. onyxi | 13 | Eastern & Western Cape | Fynbos; Grassland |
| C. orenji | 1 | Western Cape | Fynbos |
| C. parslactis | 1 | Northern Cape | Fynbos |
| C. rubrithorax | 34 | Northern & Western Cape | Forest; Fynbos |
| C. zwarti | 1 | Eastern Cape | Albany Thicket |
| Total | 62 |
Distribution maps are provided depicting the overall distribution of the genus in South Africa (Fig.


Recorded distribution for each Cryptopimpla species plotted on topographical maps. The genus is currently only recorded from the Eastern, Northern and Western Cape provinces within South Africa. Note that when two species are present in a single locality one of the species icons is larger, but centrally covered by the second species icon A C. hantami, C. kogelbergensis, C. neili, C. orenji, and C. parslactis B C. elongatus, C. fernkloofensis, C. hoerikwagga, C. onyxi, C. rubrithorax and C. zwarti.

Recorded distribution for each Cryptopimpla species plotted on biome maps. Note that when two species are present in a single locality one of the species icons is larger, but centrally covered by the second species icon A C. hantami, C. kogelbergensis, C. neili, C. orenji, and C. parslactis B C. elongatus, C. fernkloofensis, C. hoerikwagga, C. onyxi, C. rubrithorax and C. zwarti.
Digitization
The output of specimen data digitized into the Iziko South African Museum Specify 6 database is included as a supplementary excel file (Suppl. material
Identification keys
A standard dichotomous key to the African species of Cryptopimpla is presented below. Online interactive Lucid pathway and Lucid matrix keys are available on WaspWeb (
Key to African species of the genus Cryptopimpla Taschenberg, 1863
| 1 | Clypeal profile distinctly convex and bulbous (A). Pleural carinae of propodeum present, but may be weak; posterior transverse carina present and well-defined (B) | C. kogelbergensis |

|
||
| – | Clypeal profile weakly convex with a curved lip on ventral margin (a). Pleural carinae absent and posterior transverse carina of propodeum, if present, weak or reduced to a wrinkle (b) | 2 (rubrithorax species-group) |

|
||
| 2 | Metasomal tergite I punctate over most of surface (A). Median lobe of the mesocutum not raised above lateral lobes (B) | 3 |

|
||
| – | Metasomal tergite I punctate posteriorly, strigate over anterior three-quarters (a). Median lobe of the mesosocutum distinctly raised (b) | C. orenji sp. nov. |

|
||
| 3 | Mesosoma with axillar and metanotal struts subparallel, not strongly converging towards medial area (A). Propodeal anterior margin without defined medial tooth, but may have a blunt medial projection (B) | 4 |

|
||
| – | Metanotum with axillar and metonotal struts converging towards medial area (a). Propodeal anterior margin with medial tooth (b) | C. fernkloofensis |
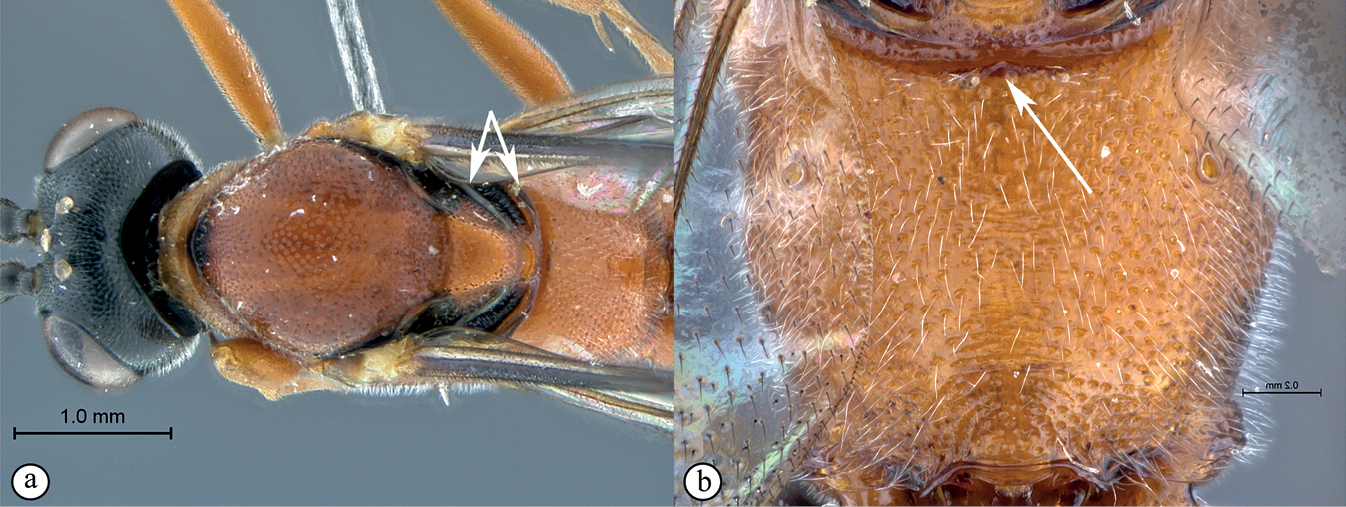
|
||
| 4 | Wings with pale microtrichia (A). Mesoscutum evenly punctate (B) | 5 |

|
||
| – | Wings with dark microtrichia, venation darker (a). Mesoscutum with fewer punctures inward of wing bases, resulting in polished areas (b) | C. parslactis |

|
||
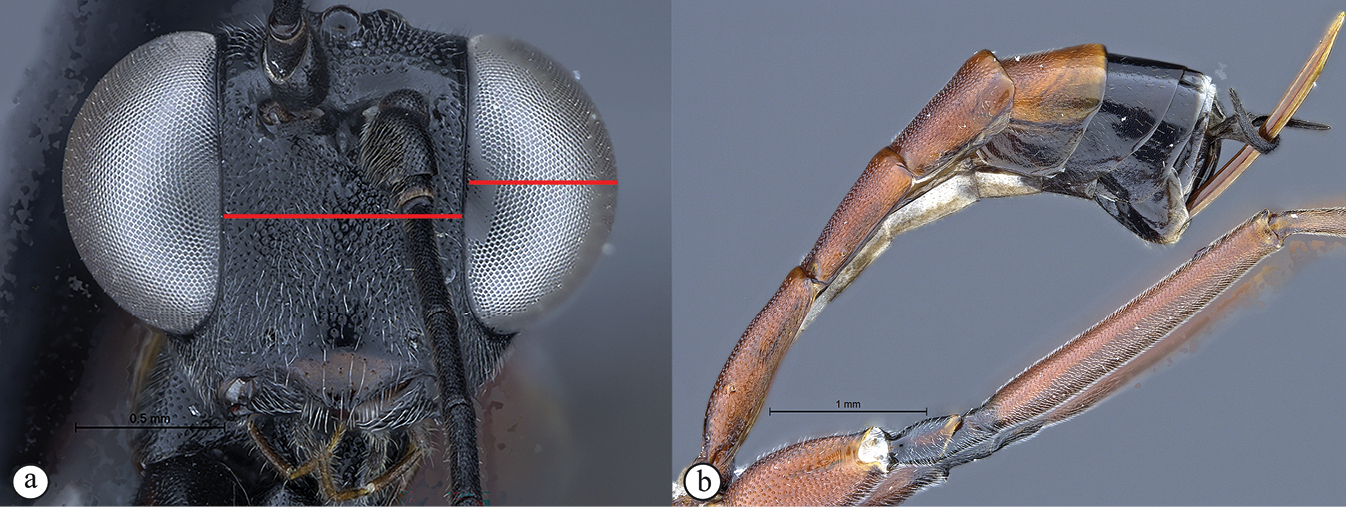
| 5 | Inter-ocular distance broad, equivalent to eye height in anterior view (A). Thyridia small and indistinct (B) | 6 |

|
||
| – | Inter-ocular distance narrow, shorter than eye height in anterior view (a). Thyridia moderately large and distinct, elongate to circular (b) | 8 |

|
||
| 6 | Male tergite IV dorso-laterally compressed (A). Posterior margin of tergite I medially projected as a blunt angle (B) | C. neili |

|
||
| – | Male tergite IV dorso-ventrally depressed (a). Posterior margin of tergite I weakly convex to straight (b) | 7 |

|
||
| 7 | Mesosoma uniformly brownish orange (A). Head and mesosoma matt, moderately covered in short setae (A). Mesopleural pit shallow with surrounding area punctate (B) | C. hantami |

|
||
| – | Mesosoma tricoloured with a distinct white pronotal collar; pronotum, propleuron, posterior mesoscutal border, ventral parts of mesopleuron and lateral areas of scutellum and metanotum black, with remaining mesosoma dark reddish brown (a). Head and mesosoma subpolished, sparsely covered in short setae (a). Mesopleural pit distinct with surrounding polished area (b) | C. hoerikwagga sp. nov. |

|
||
| 8 | Eye in anterior view narrow to moderately-sized: eye maximum width in anterior view 0.4–0.66× shortest inter-ocular distance (A). Female metasomal tergites IV–VIII slightly compressed; metasoma black (B), legs brown to black | 9 |

|
||
| – | Eye in anterior view larger, bulbous: eye maximum width in anterior view 0.73× shortest inter-ocular distance (a). Female metasoma elongated, depressed; metasoma and legs mostly rufescent (b) | C. elongatus |
|
||
| 9 | Metasomal tergite II 1.1–1.25× wider than long (A). Mesosoma predominantly rufous with some small black markings, with or without white pronotal collar (B) | 10 |

|
||
| – | Metasomal tergite II 1.09–1.25× longer than wide (a). Mesosoma black with white pronotal collar (b) | C. onyxi |

|
||
| 10 | Malar space 0.6× as long as basal mandibular width (A). Second tergite posteriorly 1.25× wider than long (B) | C. zwarti |

|
||
| – | Malar space 0.9–1.3× as long as basal mandibular width (a). Tergite II posteriorly no more than 1.1× wider than long (b) | C. rubrithorax |

|
||
Cryptopimpla
Cryptopimpla Taschenberg, 1863. Zeitschrift für die Gesammten Naturwissenschaften, 21: 292. Type-species Phytodietus blandus Gravenhorst, 1914.
Complete diagnosis
Provided in
Summary diagnosis
Afrotropical representatives of the genus can be distinguished by a combination of traits: a flagellum that tapers to a slender apex; a complete occipital carina, that joins the hypostomal carina distant from the base of the mandible; a longer upper mandibular tooth than lower tooth; absence of the epomia; a truncate-shaped fore wing areolet; the hind wing with Cu1 longer than cu-a; presence of a glymma; a strongly anteriorly narrowed first tergite, and an ovipositor that is 0.5–0.7× as long as the hind tibia (
Species-groups
The Afrotropical species cluster in two morphological species-groups:
- rubrithorax species-group (C. elongatus, C. fernkloofensis, C. hantami, C. hoerikwagga sp. nov., C. neili, C. onyxi, C. orenji sp. nov., C. parslactis, C. rubrithorax, and C. zwarti) is defined by the presence of a weakly convex clypeus with a curved lip on the ventral margin, small tentorial pits, absence of the pleural carinae, and absence of the posterior transverse carina on the propodeum.
-
kogelbergensis
species-group (C. kogelbergensis) is defined by the presence of a convex and bulbous clypeus with large tentorial pits, pleural carinae, and a distinct and well-defined posterior transverse carina on the propodeum. This species group was referred to as the
goci species-group in
Reynolds Berry and van Noort (2016) , but with the current transfer of C. goci to Lissonota Gravenhorst, 1829 in this paper the name has had to be changed to that of the single species remaining in this species-group.
Cryptopimpla hoerikwagga , sp. nov.
Type material
Holotype ♂: South Africa, W. Cape, Constantiaberge, 640 m, 34°02.5'S, 18°23.5'E, above road to mast overlooking Hout Bay, 23 Feb–2 March 1994, S. van Noort, Mesic Mountain Fynbos, Malaise trap, SAM-HYM-P00591 (SAMC).

Description
Body overall subpolished. Colour. Body mostly fulvous. Head black, clypeus and mandibles white to brown. Propleuron, fore and mid coxae, dorso-posterior margin of mesoscutum, axillary troughs of mesonotum and metanotum, submetapleural carina black. Pronotum black, pronotal collar and tegula white. Trochanters, trochantellus and tergite V brown to fulvous. Remainder of metasoma brown with tergites VII and VIII white at posterior margins. Head densely punctate. Frons unarmed. Clypeus profile weakly convex with curved lip on ventral margin. Clypeus edge convex. Upper tooth of mandible longer than lower. Setae on head and clypeus short and sparse. Tentorial pits small and indistinct. Flagellum tapered to a slender apex. Eye in lateral view 1.03 times as long as wide, maximum width in anterior view 0.55 times shortest inter-ocular distance. Mesosoma not compressed. Scuto-scutellar sulcus broad with dorso-lateral indentations. Mesoscutum evenly punctate. Epicnemial carinae present ventrally and dorsally, dorsally converging toward anterior edge of mesopleuron; mesopleural pit distinct, surrounding area polished. Propodeum without carinae, its anterior margin straight. Wings hyaline. Fore wing with two bullae close together appearing as one; vein 2m-cu sinuate; areolet truncate-shaped. Hind wing with two basal hamuli and seven distal hamuli. Metasomal tergite I with dorso-lateral wrinkles, densely punctate, with posterior margin weakly convex; tergite II 1.2 times as long as wide posteriorly, spiracle situated at anterior 0.28 of tergite (measured in lateral view), thyridia small.
CT 2.5; ML 0.9; IO 1.9; OO 1.3; Fl1 3.3; body length 9.7 mm; flagella length 9.5 mm; fore wing length 7.5 mm.
Diagnosis
This species belongs to the rubrithorax species-group and is the most strikingly coloured species of African Cryptopimpla. The mesosoma is tricoloured with a distinct white pronotal collar; a black pronotum, propleuron, posterior mesoscutal border, ventral parts of mesopleuron and lateral areas of scutellum and metanotum, with remaining mesosoma dark reddish brown. The head and mesosoma is subpolished, sparsely covered in short setae; the mesopleural pit is distinct with a surrounding polished area; axillar and metanotal struts are subparallel, not strongly converging towards the medial area; the propodeal anterior margin lacks a defined medial tooth; the metasomal tergite I is punctate over most of the surface, posterior margin weakly convex; and the thyridia are small and distinct.
Differential diagnoses
The propodeal anterior margin is straight, distinguishing the species from several members of the rubrithorax species-group (except for C. rubrithorax, C. parslactis and C. orenji) where the margin may have a blunt medial projection or medial tooth. A broad scuto-scutellar sulcus with deep lateral indentations separates the species from C. fernkloofensis, C. neili, C. hantami, C. kogelbergensis, C. parslactis, and C. orenji where the dorso-lateral indentations and/or sulcus is absent. The densely punctate tergite I distinguish this species from C. kogelbergensis and C. orenji where punctation is reduced to absent. Dorso-lateral carinae of the metasomal tergite I substituted with wrinkling separates C. hoerikwagga from C. fernkloofensis and C. neili where one or no carinae are present. Small thyridia on tergite II distinguishes this species from all other members of the rubrithorax species-group, (except for C. neili and C. hantami), where the thyridia can be elongate or moderately large and circular.
Etymology
Named after the Khoisan word for Table Mountain “hoerikwagga” which directly translates to “mountain of the sea”. Noun in apposition.
Distribution
South Africa (Western Cape) (Fig.
Comments
A rare species known only from one specimen. Intensive sampling in the type locality as well as other areas of the Cape region have so far produced no further specimens, there is, however, a major backlog of unsorted samples (
Cryptopimpla orenji , sp. nov.
Type material
Holotype ♀: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 25 April–16 May 2013, S. van Noort, Malaise trap, BH12-FYN3-M08, Burnt Mesic Mountain Fynbos, SAM-HYM-P063260 (SAMC).

Description
Body subpolished, covered in short setae. Colour. Body mostly fulvous. Epicnecium, submetapleural carinae and dorso-lateral corners of axillary troughs of meso- and metanotum black. Paraocular area of eyes, malar space, clyeus and mandibles yellow. Head densely punctate. Frons unarmed. Clypeus profile weakly convex with a curved lip on the ventral margin. Clypeus edge convex. Upper tooth of mandible longer than lower. Setae on head and clypeus short and sparse. Tentorial pits small and indistinct. Flagellum tapered to a slender apex. Eye in lateral view 1.3 times as long as wide, maximum width in anterior view 0.75 times shortest inter-ocular distance. Mesosoma not compressed. Scuto-scutellar sulcus without dorso-lateral indentations. Mesoscutum densely punctate, median lobe distinctly raised. Epicnemial carinae present ventrally and dorsally, dorsally converging toward anterior edge of mesopleuron; area surrounding mesopleural pit punctate. Propodeum with posterior transverse carinae present but weak, its anterior margin straight, spiracle elongate. Wings hyaline. Fore wing with two bullae close together appearing as one; vein 2m-cu sinuate; areolet truncate-shaped. Hind wing with two basal hamuli and six distal hamuli. Metasoma with first tergite punctate posteriorly, strigate anteriorly, with posterior margin weakly convex; tergite II of metasoma 1.8 times as long as wide posteriorly, spiracle situated at anterior 0.30 of tergite (measured in lateral view), thyridia indistinct. Tergite IV–VIII not compressed; tergite VI as wide as tergite V. Hypopygium strongly sclerotized. Ovipositor upcurved; sheath striations present.
CT 2.2; ML 0.8; IO 1.6; OO 1.6; Fl1 5; OT 0.5; body length 6.5 mm; flagella length 9.4 mm; fore wing length 6.9 mm.
Diagnosis
Cryptopimpla orenji is immediately distinguishable from all other Afrotropical Cryptopimpla by possessing a distinctly raised median lobe on the mesoscutum, and by having tergite I distinctly strigate in anterior three-quarters and only punctate posteriorly. The head coloration is fulvous; and the paraocular area of the eyes, malar space, clypeus and mandibles are yellow, a colour pattern that is unique to this species.
Differential diagnoses
The area surrounding the mesopleural pit is punctate distinguishing C. orenji from C. hoerikwagga and C. fernkloofensis where the area surrounding the pit is polished. The propodeal anterior margin is straight distinguishing the species from several members of the rubrithorax species-group (excluding C. rubrithorax, C. parslactis and C. orenji) where the margin may have a blunt medial projection or medial tooth. A scuto-scutellar sulcus without dorso-lateral indentations separates C. orenji from several closely related species (excluding C. fernkloofensis, C. parslactis and C. hoerikwagga) where the dorso-lateral indentations are present and/or the sulcus is absent. Indistinct thyridia on tergite II distinguishes the species from several members of the rubrithorax species-group (excluding C. neili, C. hantami and C. hoerikwagga) where the thyridia can be elongate to moderately large and circular.
Etymology
So named owing to the colour of this species. Orenji is the Xhosa name for orange. Noun in apposition.
Distribution
South Africa (Western Cape) (Fig.
Comments
A rare species known only from one specimen. Intensive sampling in the type locality and in other areas of the Cape region have so far produced no further specimens, there is, however, a major backlog of unsorted samples (
Lissonota goci , comb. nov.
= Cryptopimpla goci Reynolds Berry & van Noort, 2016.
Type material examined
Holotype ♂: South Africa, Western Cape, Koeberg Nature Reserve, 33°37.622'S, 18°24.259'E, 741 m, 3–31 October 1997, S. van Noort, KO97-M12, Malaise trap, West Coast Strandveld, SAM-HYM-P0474345 (SAMC).

Additional material examined for description of female
5♀: South Africa, Western Cape, Grootbos Private Nature Reserve, site LEU, 305 m, 34.531500°S, 19.482723°E, 6 Dec 2018–1 Feb 2019, S. van Noort, Malaise trap, Agulhas Limestone Fynbos, GPNR18-LEU-M09, SAM-HYM-P096893, SAM-HYM-P096967, SAM-HYM-P097347, SAM-HYM-P099594, SAM-HYM-P099621 (SAMC). 1♀: South Africa, Western Cape, Fernkloof Nature Reserve, Mosselberg, 60 m, south slope, 14 May–16 June 1995, S. van Noort, Malaise trap, Mesic Mountain Fynbos, SAM-HYM-P006315.
Additional material of males newly recorded
3♂: South Africa, W. Cape, Hermanus, Fernkloof Nature Reserve, Mosselberg, 60 m, south slope, 34°24.46'S, 19°18.00'E, 14 May–16 June 1995, S. van Noort, Malaise trap, Mesic Mountain Fynbos, SAM-HYM-P006415a-c (SAMC). 36♂: South Africa, Western Cape, Grootbos Private Nature Reserve, site LEU, 305 m, 34.531500°S, 19.482723°E, 6 Dec 2018–1 Feb 2019, S. van Noort, Malaise trap, Agulhas Limestone Fynbos, GPNR18-LEU-M09, SAM-HYM-P096887, SAM-HYM-P096888, SAM-HYM-P096892, SAM-HYM-P096895–P096899, SAM-HYM-P096901, SAM-HYM-P097300, SAM-HYM-P097305, SAM-HYM-P097307, SAM-HYM-P097335, SAM-HYM-P097336, SAM-HYM-P097340, SAM-HYM-P097341, SAM-HYM-P097346–P097348, SAM-HYM-P097351, SAM-HYM-P097353, SAM-HYM-P097394, SAM-HYM-P099598, SAM-HYM-P099617–P099620, SAM-HYM-P099622–P099624, SAM-HYM-P099626–P099631 (SAMC). 21♂: South Africa, Western Cape, Grootbos Private Nature Reserve, site LEU, 305 m, 34.531500°S, 19.482723°E, 6 Dec 2018–1 Feb 2019, S. van Noort, Malaise trap, Agulhas Limestone Fynbos, GPNR18-LEU-M14, SAM-HYM-P098708, SAM-HYM-P098715, SAM-HYM-P099730, SAM-HYM-P099734–P099737, SAM-HYM-P099741–P099744, SAM-HYM-P099745–P099747, SAM-HYM-P099749–P099751, SAM-HYM-P099753, SAM-HYM-P099754–P099756 (SAMC). 1♂: South Africa, Western Cape, Grootbos Private Nature Reserve, site FOR, 340 m, 34.54133°S, 19.43876°E, 25 Mar–31 May 2019, S. van Noort, Malaise trap, Afromontane Forest, GPNR18-FOR-M17, SAM-HYM-P099498 (SAMC). 1♂: South Africa, Western Cape, Grootbos Private Nature Reserve, site MILK, 240 m, 34.52831°S, 19.48496°E, 25 Mar–1 June 2019, S. van Noort, Malaise trap, Milkwood Scrub Forest, GPNR18-MILK-M20, SAM-HYM-P099485 (SAMC).
Description of female
Colour, sculpture and proportions as in male with the following exceptions: head with flagellum not tapered. Eye in lateral view 0.69–0.74 times as long as wide; in anterior view with maximum width slightly broader, 0.48–0.56 times shortest inter-ocular distance. Hind wing with one-two basal hamuli and seven-eight distal hamuli. Metasoma with tergite I impunctate, wrinkles or a single carina present dorso-laterally with no secondary carina leading from the single carina to the spiracle; second tergite 0.98–1.39 times longer than broad, spiracle situated more anteriorly at 0.27–0.3 of tergite (measured in lateral view), ovipositor 2.31–2.36 times longer than hind tibia.
Body length 7.1–8.7 mm; antenna length 7.1–8.1 mm; fore wing length 5.3–6.1 mm.
Distribution
South Africa (Western Cape) (Fig.
Cryptopimpla elongatus
Type material examined
Holotype ♀: South Africa, Northern Cape, Hantam National Botanical Garden, 31°24.274'S, 19°09.164'E, 755 m, 22 May–12 June 2008, S. van Noort, GL07-DOL1-M39, Malaise trap, Nieuwoudtville-Roggeveld Dolerite Renosterveld, SAM-HYM-P047468 (SAMC).
Cryptopimpla fernkloofensis
Type material examined
Holotype ♂: South Africa, Western Cape, Fernkloof Nature Reserve, 33°39.941'S, 21°53.505'E, 300–340 m, 13 May 1995, S. van Noort, Sweep, Mesic Mountain Fynbos, SAM-HYM-P008237 (SAMC).
Cryptopimpla hantami
Type material examined
Holotype ♀: South Africa, Northern Cape, Hantam National Botanical Garden, 31°24.182'S, 19°08.587'E, 741 m, 17 March–21 April 2008, S. van Noort, GL07-REN3-M24, Malaise trap, Nieuwoudtville Shale Renosterveld, SAM-HYM-P047467 (SAMC).
Cryptopimpla kogelbergensis
Type material examined
Holotype ♀: South Africa, Western Cape, Kogelberg Nature Reserve, 34°16.481'S, 19°01.033'E, 118 m, 16 May–16 June 1999, S. van Noort, KO98-M23, Malaise trap, Mesic Mountain Fynbos, last burnt c. 1988, SAM-HYM-P047475 (SAMC).
Additional material newly recorded
1♀: South Africa, Northern Cape, Hantam National Botanical Garden, 31°24.274'S, 19°09.164'E, 23 March–06 May 2008, S. van Noort, GL07-REN3-M38, 741 m, Malaise trap, Nieuwoudtville-Roggeveld Dolerite Renosterveld, SAM-HYM-P064320 (SAMC).
Cryptopimpla neili
Type material examined
Holotype ♂: South Africa, Western Cape, Kogelberg Nature Reserve, 34°16.481'S, 19°01.033'E, 118 m, 16 March 1999–16 April 1999, S. van Noort, KO98-M18, Malaise trap, Mesic Mountain Fynbos, last burnt c. 1988, SAM-HYM-P047436 (SAMC).
Cryptopimpla onyxi
Type material examined
Holotype ♀: South Africa, Western Cape, Walker Bay Nature Reserve, 34°27.414'S, 19°21.393'E, 57 m, 14 May–14 June 1997, S. van Noort, WB97-M01, Malaise trap, South coast Strandveld, SAM-HYM-P047460 (SAMC).
Additional material newly recorded
1♂: South Africa, Western Cape, Grootbos Private Nature Reserve, site LEU, 305 m, 34.531500°S, 19.482723°E, 25 March–31 May 2019, S. van Noort, Malaise trap, Agulhas Limestone Fynbos, GPNR18-LEU-M19, SAM-HYM-P098730 (SAMC). 1♂: South Africa, Western Cape, Grootbos Private Nature Reserve, site MILK, 240 m, 34.52831°S, 19.48496°E, 1 June–7 Aug 2019, S. van Noort, Malaise trap, Milkwood Scrub Forest, GPNR18-LEU-M24, SAM-HYM-P101469 (SAMC). 1♀: South Africa, Eastern Cape, Winterberg, The Hoek farm, 31°21.260'S, 26°23.001'E, 1879 m, 6.x.2010–18.i.2011, S. van Noort, Malaise trap, Amathole Mistbelt Grassland, WTB09-GRA1-M05, SAM-HYM-P062421 (SAMC).
Cryptopimpla parslactis
Type material examined
Holotype ♂: South Africa, Northern Cape, Hantam National Botanical Garden, 31°23.802'S, 19°08.799'E, 752 m, 23 July–23 Aug 2008, S. van Noort, GL07-REN1-M43, Malaise trap, Nieuwoudtville Shale Renosterveld, SAM-HYM-P044547 (SAMC).
Cryptopimpla rubrithorax
Type material examined
Holotype ♀: South Africa, Western Cape, Elsenberg, 11 October 1914, Mally and Petty, SAM-HYM-P000874 (SAMC).
Additional material newly recorded
3♂: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 28 Aug–28 Sept 2012, S. van Noort, Malaise trap, Mesic Mountain Fynbos, BH12-FYN3-M02, SAM-HYM-P063982, SAM-HYM-P064071, SAM-HYM-P097386 (SAMC). 4♀,3♂: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 3–25 April 2013, S. van Noort, Malaise trap, Burnt Mesic Mountain Fynbos, BH12-FYN3-M03, SAM-HYM-P063159, SAM-HYM-P063497, SAM-HYM-P093877–P093878, SAM-HYM-P093879–P093881 (SAMC). 3♂: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 28 Sept–24 Oct 2012, S. van Noort, Malaise trap, Mesic Mountain Fynbos, BH12-FYN3-M07, SAM-HYM-P063492, SAM-HYM-P063537 (SAMC). 3♂: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 8 Aug–2 Oct 2013, S. van Noort, Malaise trap, Burnt Mesic Mountain Fynbos, BH12-FYN3-M12, SAM-HYM-P063516, SAM-HYM-P063704, SAM-HYM-P063716 (SAMC). 3♀,1♂: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 25 April–16 May 2013, S. van Noort, Malaise trap, Burnt Mesic Mountain Fynbos, BH12-FYN3-M08, SAM-HYM-P063076, SAM-HYM-P063278, SAM-HYM-P093875–P093876 (SAMC). 2♀: South Africa, W. Cape, Koeberg Nature Reserve, 33°37.622'S, 18°24.259'E, 8 Aug–5 Sept 1997, S. van Noort, Malaise trap, KO97-M07, West Coast Strandveld, SAM-HYM-P047476 (SAMC). 1♀: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 2–22 October 2013, S. van Noort, Malaise trap, Mesic Mountain Fynbos, BH12-FYN3-M13, SAM-HYM-P064020 (SAMC). 1♀,1♂: South Africa, Western Cape, Banghoek Valley, Grootbos Private Nature Reserve, site LEU, 305 m, 34.531500°S, 19.482723°E, 25 March–31 May 2019, S. van Noort, Malaise trap, Agulhas Limestone Fynbos, GPNR18-LEU-M19, SAM-HYM-P098731, SAM-HYM-P098765 (SAMC). 1♀: South Africa, Western Cape, Banghoek Valley, Dwarsriviershoek Farm, 33°56.232'S, 18°57.711'E, 410 m, 24 Oct–10 Dec 2012, S. van Noort, Malaise trap, Mesic Mountain Fynbos, BH12-FYN3-M04, SAM-HYM-P064919 (SAMC). 1♀: South Africa, Western Cape, Table Mountain National Park, Orangekloof, Disa River, 34°0.035'S, 18°23.492'E, 136 m, 11 Nov–11 Dec 2014, S. van Noort, Malaise trap, Afromontane Forest, OGK13-FOR1-M27, SAM-HYM-P062973 (SAMC).
Cryptopimpla zwarti
Type material examined
Holotype ♀: South Africa, Eastern Cape, Grahamstown, Faraway Farm 33°19'S, 19°26.31'E, April 1990, I. Crampton, Malaise trap, SAM-HYM-P005220 (SAMC).
Discussion
Species-groups
The two newly described species (C. orenji sp. nov. and C. hoerikwagga sp. nov.) both belong to the rubrithorax species-group. In addition to possessing the morphological characters that distinguish members of the rubrithorax species-group, these two new species also have larger fore wings (length = 5.8–7.5 mm) compared to the kogelbergensis species-group where the fore wing lengths are smaller (length = 4.6–5.1 mm). Cryptopimpla hoerikwagga sp. nov. is the largest African Cryptopimpla species (body length 9.7 mm; fore wing length 7.5 mm) and C. kogelbergensis is the smallest African Cryptopimpla species (body length 4.2–5.6 mm; fore wing length 4.6–5.1 mm). The remaining species have sizes ranging between these two extremes (body length 6.5–9.4 mm; fore wing length 5.8–7.2 mm).
The maximum length of the ovipositor sheath relative to the hind tibia for the genus ranges from 0.7× for Afrotropical species (
The discovery of the female of Lissonota goci comb. nov. has allowed us to reassess generic affinities of this species and based on female characters this species is better placed within Lissonota. The apical 0.3–0.4 portion of the flagellum is tapered to a slender apex in Cryptopimpla whereas the female flagellum in this species, as typical for Lissonota, is only weakly tapered at the apex (
Distribution and diversity
Cryptopimpla hoerikwagga sp. nov. and C. orenji sp. nov. are described based on single specimens that were collected from the western slopes of the Constantiaberg mountain, and Banghoek Valley adjacent to the Helshoogte mountain, of Western Cape South Africa, respectively. These are areas with no previous records for Cryptopimpla. Both have been collected in Mesic Mountain Fynbos, a vegetation type that is a habitat association for most of the previously described species (i.e. C. fernkloofensis, C. neili, C. onyxi, C. rubrithorax and C. kogelbergensis). Approximately 55% of Afrotropical Cryptopimpla species have been described based on a single specimen, and 90% of the overall Afrotropical Cryptopimpla species diversity is currently recorded from fynbos (Fig.
Due to the relatively limited availability of specimens for several species within Cryptopimpla, any assessments of the distribution and diversification of the different species are still likely to be biased. This is corroborated by unique locality records for the newly described species presented in this paper. While sustained continuous inventory surveys over the last three decades has revealed the genus to be increasingly species rich, it is still rare in terms of abundance with more than half of the Afrotropical species represented by a single specimen. Further specimens will no doubt be recovered from the backlog of unsorted samples resulting from 31 years of continuous inventory surveys run by Simon van Noort, using a range of collecting methods (Malaise traps, yellow pan traps, yellow funnel traps, pitfall traps, sweeping, Winkler bag extraction of leaf litter and UV light trapping) (
Cryptopimpla rubrithorax is the most common Afrotropical species, occurring across various vegetation types within the Cape Floristic Region, including Strandveld, Mesic Mountain Fynbos, Agulhas Limestone Fynbos, Renosterveld, and Afromontane forest (
Acknowledgements
Cape Nature, the Eastern Cape Department of Environmental Affairs, and the Northern Cape Department of Nature and Environmental Conservation provided collecting permits. SANParks provided a research permit. SvN was funded by South African NRF (National Research Foundation) grants: GUN 2068865; GUN 61497; GUN 79004; GUN 79211; GUN 81139. Part of the South African field work conducted by SvN was funded by the National Science Foundation under PlatyPBI grant No. DEB–0614764 to N.F. Johnson and A.D. Austin. Thanks to Gavin Broad and Niklas Johansson for their valuable reviews of the manuscript.
Project conceptualization (SvN, TR); funding (SvN); field work (SvN); data curation (TR, SvN); taxonomy – species descriptions and species delimitation assessments (TR, SvN); imaging (TR); figure plates and identification key plate production (TR, SvN); production of identification keys (TR, SvN); manuscript production and editing (TR, SvN).
References
- Broad GR, Shaw MR, Fitton MG (2018) Ichneumonid wasps (Hymenoptera: Ichneumonidae): their classification and biology. Handbooks for the Identification of British Insects 7. Royal Entomological Society, 418 pp.
- Broad GR (2022) Presentation: Large-scale species revisions. Darwin wasp Conference, Öland. July 2022.
- Brock JP (2017) The banchine wasps (Ichneumonidae: Banchinae) of the British Isles. Handbooks for the identification of British insects 7. Royal Entomological Society, 150 pp.
- Cardoso P, Erwin T, Borges P, New T (2011) The seven impediments in invertebrate conservation and how to overcome them. Biological Conservation 144: 2647–2655. https://doi.org/10.1016/j.biocon.2011.07.024
- Cardoso P, Leather SR (2019) Predicting a global insect apocalypse. Insect Conservation and Diversity 12: 263–267. https://doi.org/10.1111/icad.12367
- Cardoso P, Barton PS, Birkhofer K, Chichorro F, Deacon C, Fartmann T, Fukushima CS, Gaigher R, Habel JC, Hallmann CA, Hill MJ, Hochkirch A, Kwak ML, Mammola S, Noriega JA, Orfinger AB, Pedraza F, Pryke JS, Roque FO, Settele J, Samways MJ (2020) Scientists’ warning to humanity on insect extinctions. Biological Conservation 242: e108426. https://doi.org/10.1016/j.biocon.2020.108426
- Ceballos G, Ehrlich PR, Barnosky AD, García A, Pringle RM, Palmer TM (2015) Accelerated modern human-induced species losses: Entering the sixth mass extinction. Science Advances 1(5): e1400253. https://doi.org/10.1126/sciadv.1400253
- Gauld ID (1991) The Ichneumonidae of Costa Rica 1. Memoirs of the American Entomological Institute 47: 1–589.
- Gauld ID, Gomez J, Godoy C (2002) Subfamily Banchinae. In: Gauld ID (Ed.) The Ichneumonidae of Costa Rica, 4. Memoirs of the American Entomological Institute 66: 263–746.
- Habel J, Schmitt T, Gros P, Ulrich W (2023) Breakpoints in butterfly decline in Central Europe over the last century. Science of The Total Environment 851: e158315. https://doi.org/10.1016/j.scitotenv.2022.158315
- Holmgren AE (1860) Forsok till uppstallning och beskrifning af Sveriges Ichneumonider. Tredje Serien. Fam. Pimplariae. (Monographia Pimplariarum Sueciae). Kongliga Svenska Vetenskapsakademiens Handlingar 3: 1–76.
- Kang GW, Kolarov J, Lee JW (2019) Cryptopimpla (Hymenoptera, Ichneumonidae, Banchinae) of South Korea, with description of two new species. ZooKeys 830: 99–109. https://doi.org/10.3897/zookeys.830.31974
- Kuslitzky WS (2007) 12. Subfamily Banchinae. In: Lelej AS (Ed.) A Key to the Insects of Russian Far East (Vol. 4). Neuropteroidea, Mecoptera, Hymenoptera. Part 5.: Dal’nauka, Vladivostok 433–472. [in Russian]
- Linder HP (2003) The radiation of the Cape flora, southern Africa. Biological Reviews 78: 597–638. https://doi.org/10.1017/S1464793103006171
- Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403(6772): 853–858. https://doi.org/10.1038/35002501
- Momoi S, Sugawara H, Honma K (1975) Ichneumonid and Braconid parasites of Lepidopterous leaf-rollers of economic importance in horticulture and teaculture. In: Yasumatsu K, Mori H (Eds) Approaches to Biological Control 7: 47–60.
- Penev L, Sharkey M, Erwin T, van Noort S, Buffington M, Seltmann K, Johnson N, Taylor M, Thompson C, Dallwitz M (2009) Data publication and dissemination of interactive keys under the open access model. ZooKeys 21: 1–17. https://doi.org/10.3897/zookeys.21.274
- Quicke DLJ (2015) The braconid and ichneumonid parasitoid wasps: Biology, systematics, evolution and ecology. Wiley Blackwell: 1–704. https://doi.org/10.1002/9781118907085
- Reynolds Berry T (2019) Systematics of the parasitoid wasp subfamily Banchinae (Hymenoptera; Ichneumonidae) in the Afrotropical region. PhD thesis. Stellenbosch University (South Africa). http://hdl.handle.net/10019.1/106917
- Reynolds Berry T, van Noort S (2016) Review of Afrotropical Cryptopimpla Taschenberg (Hymenoptera, Ichneumonidae, Banchinae), with description of nine new species. Zookeys 640: 103–137. https://doi.org/10.3897/zookeys.640.10334
- Reynolds Berry T, van Noort S (2020) Revision of the endemic Afrotropical genus Tetractenion (Hymenoptera, Ichneumonidae) with an identification key to genera of Banchinae for the region. ZooKeys 1007: 49–84. https://doi.org/10.3897/zookeys.1007.55543
- Sharkey M, Yu D, van Noort S, Seltmann K, Penev L (2009) Revision of the Oriental genera of Agathidinae (Hymenoptera, Braconidae) with an emphasis on Thailand and interactive keys to genera published in three different formats. ZooKeys 21: 19–54. https://doi.org/10.3897/zookeys.21.271
- Shaw M (2006) Habitat considerations for parasitic wasps (Hymenoptera). Journal of Insect Conservation 10: 117–127. https://doi.org/10.1007/s10841-006-6288-1
- Shaw MR (2022) Host ranges of Aleiodes species (Hymenoptera: Braconidae), and an evolutionary hypothesis. In: Melika G, Thuróczy C (Eds) Parasitic Wasps: Evolution, Systematics, Biodiversity and Biological Control. Agroinform, Budapest, 321–327.
- Sheng ML (2011) Five new species of the genus Cryptopimpla Taschenberg (Hymenoptera, Ichneumonidae) with a key to species known from China. ZooKeys 117: 9–49. https://doi.org/10.3897/zookeys.117.1302
- Sheng ML, Zheng H (2005) The genus Cryptopimpla from China (Hymenoptera, Ichneumonidae). Acta Zootaxonomica Sinica 30: 415–418.
- Shorthouse DP (2010) SimpleMappr, an online tool to produce publication-quality point maps. https://www.simplemappr.net [Accessed 2022/07/28]
- Snijman D, Perry P (1987) A floristic analysis of the Nieuwoudtville Wild Flower Reserve, north-western Cape. South African Journal of Botany 53: 445–454. https://doi.org/10.1016/S0254-6299(16)31378-3
- Takasuka K, Watanabe K, Konishi K (2011) Genus Cryptopimpla Taschenberg new to Sulawesi, Indonesia, with description of a new species (Hymenoptera, Ichneumonidae, Banchinae). Journal of Hymenoptera Research 23: 65–75. https://doi.org/10.3897/jhr.23.1595
- Taschenberg EL (1863) Die Schlupfwespenfamilie Pimplariae der deutschen Fauna, mit besonderer Rücksicht auf die Umgegend von Halle. Zeitschrift für die Gesammten Naturwissenschaften 21: 245–305.
- Topp EN, Loos J (2019) Fragmented landscape, fragmented knowledge: a synthesis of renosterveld ecology and conservation. Environmental Conservation 46: 171–179. https://doi.org/10.1017/S0376892918000498
- Townes HK (1969) Genera of Ichneumonidae, Part 3 (Lycorininae, Banchinae, Scolobatinae, Porizontinae). Memoirs of the American Entomological Institute 13: 1–307.
- Townes HK, Townes M (1978) Ichneumon-flies of America north of Mexico: 7. Subfamily Banchinae, tribes Lissonotini and Banchini. Memoirs of the American Entomological Institute 26: 1–614.
- van Noort S (2019) Assessing the status quo of Afrotropical ichneumonid knowledge. Conference: identifying the next challenges in ichneumonid systematics and evolutionary ecology. Basel, Sweden, 24–29 June 2019. https://doi.org/10.5281/zenodo.3395821
- van Noort S (2022) Keynote presentation: Darwin wasps for Africa: Status quo of Afrotropical Ichneumonidae. #HYMATHON2022 A 24-hour virtual symposium from the International Society of Hymenopterists. 31 March–1 April 2022.
- van Noort S (2023a) WaspWeb: Hymenoptera of the Afrotropical region. www.waspweb.org [Accessed on 24 March 2023]
- van Noort S (2023b) Thirty years of sampling effort and status quo of African Darwin wasp diversity (Ichneumonidae, Hymenoptera). 23rd Congress of the Entomological Society of Southern Africa (ESSA), 12–14 July 2023, Stellenbosch University, Stellenbosch, South Africa.
- van Wyk AE, Smith GF (2001) Regions of floristic endemism in southern Africa: A review with emphasis on succulents. Umdaus Press (Hatfield, South Africa), 199 pp.
- Von Hase A, Rouget M, Maze K, Helme N (2003) A fine-scale conservation plan for Cape lowlands renosterveld: technical report. Report CCU [www document]. http://bgis.sanbi.org/clr
- Wahl DB, Sharkey MJ (1993) Chapter 10. Superfamily Ichneumonoidea. In: Goulet H, Huber JT (Eds) Hymenoptera of the World: An Identification Guide to Families. Agriculture Canada, 358–509.
- Yu DSK, van Achterberg C, Hortsmann K (2016) Taxapad 2016, Ichneumonidae 2011. www.taxapad.com [Accessed on 30/11/2018]
Supplementary materials
The output of Cryptopimpla (Ichneumonidae, Hymenoptera) specimen data digitized into the Iziko South African Museum Specify 6 database provided as a supplementary excel file
Data type: xls
Lucid Interchange Format version 3 (LIF3) files to the WaspWeb online Lucid matrix identification key to Afrotropical species of Cryptopimpla (Ichneumonidae, Hymenoptera)
Data type: lif3
Explanation note: The LIF3 file is an XML-based file that stores all the Lucid3 key data, allowing exchange of the key with other key developers.