





(C) 2012 Juan José Martínez. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Pambolus albospina sp. n. and Pambolus rastafari sp. n. are described from Jamaica. Pambolus rastafari can be distinguished from all other Neotropical species of the genus by its antennal color pattern and the smooth sculpture of the head and mesoscutum. Pambolus albospina is morphologically close to Pambolus hemitaeniatus van Achterberg, from which it can be distinguished by the number of white antennal segments and the sculpture on the first metasomal tergite. These two new species constitute the first records of the genus for the Caribbean Islands.
Caribbean, Hormiinae, Pambolinae, Taxonomy
Pambolus Haliday is a braconid genus with 42 currently valid species from all biogeographic regions (
In the Neotropical Region, Pambolus includes 12 described mainland species that are distributed from Honduras to northern Argentina (
Specimens were collected using yellow pan traps at two localities in Jamaica.
Morphological terminology follows
Specimens are deposited at Colección Nacional de Insectos (CNIN), Instituto de Biología, Universidad Nacional Autónoma de México, and at Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina (MACN).
Resultsurn:lsid:zoobank.org:act:19BA7908-B656-4B6E-A661-F9C68582B370
http://species-id.net/wiki/Pambolus_albospina
Figures 1–6Following the key to Neotropical species (
Female. Color: Mostly reddish brown (fig. 1); scape and base of pedicel pale yellow, apex of pedicel and first five flagellomeres honey yellow, sixth to eleventh flagellomeres gradually darkening from brownish yellow to black, further flagellomeres (12th to 24th ) entirely white. Palpi white; lower area of temple, gena, face, clypeus and labrum reddish brown; frons, vertex and upper area of temple orange brown, ocellar triangle darkened. Mesoscutum, scutellum and propodeum except propodeal spines, orange brown; pronotum, mesopleuron and metapleuron darker, rusty brown; propodeal spines entirely white. First metasomal tergite reddish brown, reminder of metasoma brown. Coxae, trochanters and trochatelli white, femora white basally and brown on apical third fourth, tibiae whitish, tarsi light brown. Fore wing with faint infuscate bands, hind wing hyaline, pterostigma and veins brown, except vein r-m and junction of veins 2RS and 2M, which are not pigmented.
Head: Antenna (fig. 5) with 24 flagellomeres, first flagellomere 1.1 times longer than second, first and second flagellomeres 5.5 and 5.0 times longer than their maximum width respectively. Scapus oblique apically; in dorsal view (fig. 3) length of eyes 1.9 times temple; POL 1.5 times OD and 0.7 times OOL; in dorsal view width of head 1.7 times its maximum length; face flattened and clypeus weakly convex in lateral view; face (fig. 2) weakly coriaceous near toruli, turning smooth near clypeus; clypeus largely smooth; anterior tentorial pit big and circular; frons, vertex and temple rugulose; gena smooth and shining (fig. 4); malar space 1.5 times basal width of mandible; occipital carina complete, meeting hypostomal carina ventrally.
Mesosoma: Length of mesosoma 1.4 times its maximum height and 1.6 times its maximum width; mesosoma setose; pronotum largely smooth, pronotal furrow deep and scrobiculate; propleuron convex and weakly coriaceoous; mesopleuron smooth (fig. 4), precoxal sulcus deep and scrobiculate, occupying two thirds length of mesopleuron, posterior mesopleural furrow strongly scrobiculate; subalar area rugose; episternal scrobe deep; prepectal and postpectal carinae strongly developed; metapleuron reticualte; mesoscutum (fig. 3) acinose-coriaceous; notauli wide and scrobiculate anteriorly, rather obscured in a median area posteriorly; scutellum coriaceous and slightly convex; scutellar sulcus with five carinae, its median length 0.6 times as long as scutellum; propodeum areolate rugose, with a long and narrow areola medially (fig. 3), areola irregularly rugose anteriorly and with four transverse carinae posteriorly, spines of propodeum 0.8 times as long as fore basitarsus.
Wings: Wings (fig. 6) densely setose. Fore wing: veins r and 3RSa basally widened; r:3RSa:3RSb = 2:5:16; 2RS:2M:r-m = 6:10:3; vein (RS+M)a straight; vein cu-a just interstitial and almost indistinct; first subdiscal cell open. Hind wing: vein cu-a oblique, directed towards wing base; M+CU:1-M = 2:5.
Legs: Hind coxa unsculptured dorsally; length of femur, tibia and basitarsus of hind leg 4.4, 7.6 and 6.0 times their width, respectively; hind femur very weakly coriaceous.
Metasoma: Length of first tergite 0.7 times its apical width, its median area well delimited by carinae but entirely smooth, lateral areas also smooth (fig. 3); second metasomal tergum with a few basal striations, otherwise smooth, remaining terga completely smooth; combined length of second and third tergites as long as their maximum width; length of ovipositor sheath 0.4 times the length of metasoma.
Male Similar to female, except for its slightly lighter body color and its much longer antennae, with 31 flagellomeres, five apical flagellomeres entirely white.

Pambolus albospina sp. n. 1 Habitus of female (lateral view) 2 head (anterior view) 3 head, mesosoma and first metasomal segment (dorsal view) 4 head, mesosoma and first metasomal segment (lateral view) 5 antenna (dorsal view) 6 wings.
The specific epithet refers to the white propodeal spines of this species.
Holotype female: Jamaica, Trelawny, Windsor, 18.35752, -76.66406, 82m, 19–21.xi.2010, yellow pan traps, F. S. Ceccarelli col. GenBank accession no. JQ268751. (CNIN IB-UNAM) Paratypes: three males, Jamaica, Saint Andrew, New Castle, 18.06840, -76.7119, 860m, 11.xi.2010, F. S. Ceccarelli col. (CNIN IB-UNAM, MACN); GenBank accession no. JQ268752.
urn:lsid:zoobank.org:act:76DBA72C-1920-4BF3-860E-860B88A1739F
http://species-id.net/wiki/Pambolus_rastafari
Figures 7– 15Following the keys to Neotropical species of Pambolus (
Female. Color: Mostly chestnut brown (fig. 7); palpi, pedicel, second flagellomere, coxae, trochanters and trochantelli, all femora and all tibiae basally ivory white; flagellomeres beyond 12th and fore tarsus light brown; annellus of first flagellomere and apex of all trochantelli distinctly orange-brown; wings almost entirely infuscate, with subhyaline areas at junction of veins 2RS and 2M and along vein r-m; remaining veins light brown.
Head: Antenna broken, 13 flagellomeres remaining, first flagellomere 1.2 times longer than second, first and second flagellomeres 5.0 and 3.9 times longer than their maximum width, respectively. Scapus oblique apically. In dorsal view (fig. 9) length of eyes 1.9 times the length of temple; POL as long as OD and 0.4 times OOL; in dorsal view width of head 0.6 times its median length; face rather flattened and clypeus weakly convex in lateral view (fig. 10); face smooth and setose (fig. 8); clypeus smooth; anterior tentorial pit big and circular; frons smooth and slightly concave; vertex, temple and gena also smooth and shining; malar space 2.0 times basal width of mandible; occipital carina complete, strongly developed, meeting hypostomal carina ventrally.
Mesosoma: Length of mesosoma 1.4 times its maximum height and 1.6-1.7 times its maximum width; mesosoma setose; pronotum largely smooth, slightly coriaceous anteriorly, pronotal furrow present only medially and scrobiculate; propleuron convex and smooth; mesopleuron smooth (fig. 11), precoxal sulcus deep and scrobiculate, occupying two thirds the length of the mesopleuron, subalar and posterior mesopleural furrows scrobiculate; prepectal and postpectal carinae strongly developed; metapleuron irregularly rugose; mesoscutum smooth and shining (fig. 12); notauli wide and crenulate on anterior edge of mesoscutum, indistinct posteriorly; mesoscutum with a median V-shaped posterior pit; scutellar disc smooth; scutellar sulcus with five coarse and oblique carinae, its median length 0.5 times as long as scutellar disc; propodeum (fig. 13) areolate rugose, with a long and narrow areola medially, areola with a median longitudinal carina anteriorly and with three transverse carinae posteriorly, spines of propodeum 0.7 times as long as fore basitarsus.
Wings: Wings densely setose. Fore wing (fig. 14): veins r and 3-SR basally widened; r:3RSa:3RSb = 3:8:12; 2RS:2M:r-m = 9:17:5; veins (RS+M)b and r-m unpigmented; vein (RS+M)a straight; vein cu-a insterstitial; first subdiscal cell open. Hind wing (fig. 15): vein cu-a vertical; M+CU:1-M = 2:5.
Legs: Hind coxa largely finely striate dorsally; length of femur, tibia and basitarsus of hind leg 4.4, 8.4 and 5.3 times their width, respectively; hind femur very weakly coriaceous.
Metasoma: Length of first tergite 0.8 times its apical width, its median area wide and coarsely longitudinally striate (fig. 13), lateral areas smooth; remaining terga completely smooth and shining; combined length of second and third tergites as long as their maximum width; length of ovipositor sheath 0.4-0.5 times the length of metasoma.
Male. Unknown.

Pambolus rastafari sp. n. 7 habitus of female (lateral view) 8 head (anterior view) 9 head (dorsal view) 10 head and basal antennomeres (lateral view) 11 mesosoma (lateral view) 12 mesoscutum and sctuellum (dorsal view) 13 propodeum and first metasomal segment (dorsal view).
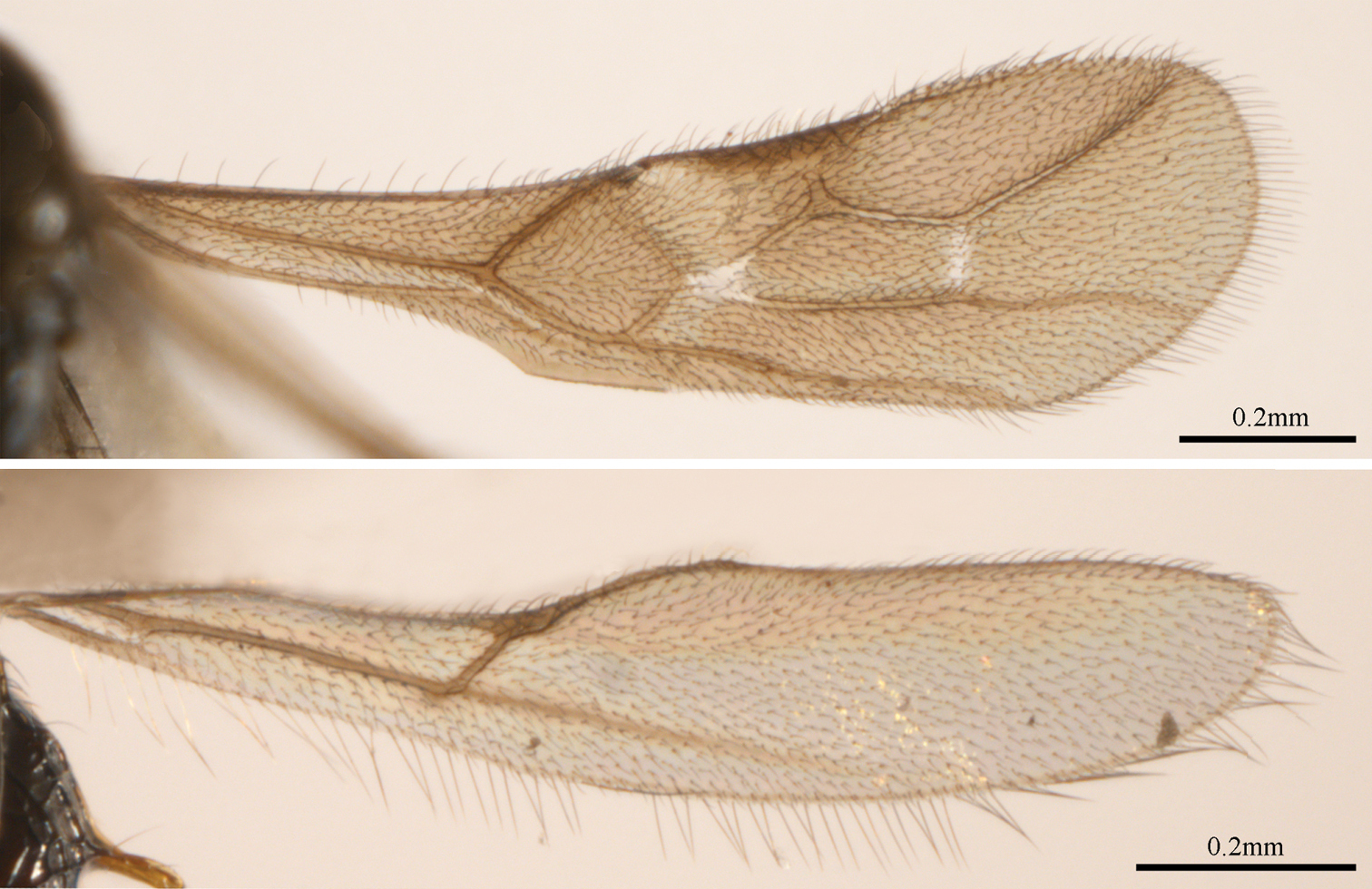
Pambolus rastafari sp. n. 14 fore wing 15 hind wing.
The specific epithet refers to the religious and social movement from Jamaica.
Holotype female: Jamaica, Saint Andrew, New Castle, 18.06840, -76.7119, 860m, 11.xi.2010, F. S. Ceccarelli col. (CNIN IB-UNAM); GenBank accession no. JQ268750. Paratype: one female, same data as holotype (MACN).
Grateful thanks are extended to Susana Guzmán Gómez (UNIBIO, IB- UNAM, Mexico), for her assistance in taking the digital images. To Dwight Robinson from UWI Mona, Kingston, Jamaica, for his assistance during our collecting trip. We also thank two anonymous reviewers for their helpful comments on an earlier version of the manuscript. This work was supported by postdoctoral fellowships given by the Universidad Nacional Autónoma de México (UNAM; DGAPA fellowships) to JJM, FSC and SFC, and by grants given by the Consejo Nacional de Ciencia y Tecnología [proyecto Ciencia Básica CONACyT no. 511 and Programa de Cooperación Bilateral México-Argentina (CONACyT-CONICET)] and by the Ministerio de Ciencia e Innovación (CGL2010-15786; Spain) to AZR.