





(C) 2012 Juliano F. Nunes. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
We redescribe the poorly known, gall associated doryctine wasp Mononeuron duguetiae Fischer (Braconidae) based on museum and recently collected material. This species shares various morphological features with the members of the also gall associated Allorhogas Gahan, suggesting their close relationship. A Bayesian phylogenetic analysis using mitochondrial (COI) and nuclear (28S) markers recovered Mononeuron duguetiae within a clade with another five gall associated doryctine genera + Heterospilus Haliday. Surprisingly, Mononeuron duguetiae was recovered, though with low support, as the sister taxon of a Monitoriella Hedqvist + Labania Hedqvist clade, and not closely related to Allorhogas. Information is provided about the galls on Duguetia furfuracea (St. Hil.) (Annonaceae) where the specimens of Mononeuron duguetiae were reared.
redescription, Braconidae, Doryctinae, gall associated, Mononeuron
The braconid subfamily Doryctinae is a cosmopolitan, highly diverse group of parasitoid wasps containing more than 1, 300 described species (
The doryctine genus Mononeuron Fischer was erected by
Here, we redescribe Mononeuron duguetiae based on museum (including the holotype specimen) and recently collected material, and provide information about the galls where the species was reared. We also employed one mitochondrial (mt) [cytochrome oxidase I (COI)] and one nuclear [28S ribosomal (r) RNA gene] marker to elucidate the phylogenetic affinities of this taxon.
MethodsAll the specimens examined in this study were collected in the type locality of the species, the vegetation of which consists of the remains of Brazilian savannah (‘cerrado’) forest owned by the Universidade Federal de São Carlos, in São Carlos, São Paulo, Brazil. Some of the specimens were reared from galls on Duguetia furfuracea in February of 2011. The specimens were preserved in 100% ethanol and kept at -20°C until they were mounted or processed for DNA sequencing. The examined material is deposited in the Departamento de Ecologia e Biologia Evolutiva, Universidade Federal de São Carlos, São Carlos, SP, Brazil (DCBU), the Colección Nacional de Insectos, Instituto de Biología, Universidad Nacional Atónoma de México (CNIN IB-UNAM), and the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina (MACN).
The terminology employed follows
The phylogenetic placement of Mononeuron was assessed using DNA sequences of one fast evolving (629 bp of the COI mtDNA gene) and one slow evolving [~650 bp of the second and third domain regions of the 28S rRNA gene] gene marker. Two COI and one 28S sequence were generated from specimens of Mononeuron duguetiae (COI: GenBank accession nos JQ320372-3; 28S: GenBank accession no. JQ320371). Protocols for genomic DNA extraction of the samples (using a non-destructive technique) and amplification of the PCR products were the same as those employed by
COI genetic distances between the two sequenced specimens of Mononeuron duguetiae and among these and previously published sequences of species belonging to Heterospathius, Heterospilus and to the gall associated doryctine genera Allorhogas Gahan, Labania Hedqvist, Monitoriella Hedqvist, Percnobracon Kieffer & Jörgensen and Psenobolus Reinhard were calculated using the K2P distance model (
A Bayesian MCMC partitioned analysis was run with MrBayes version 3.1.2 (
We used a Bayesian approach (
Corrected COI genetic distances between the two specimens of Mononeuron dugetiae and among the latter specimens and the species belonging to other gall associated genera were of 0.5% (3 bp) and of 13.5 to 16.5% (71 to 86 bp), respectively. Corrected COI distances among species of the above gall associated taxa and species of Heterospilus varied from 14.4 to 21.1 % (79 to 111 bp). Our Bayesian phylogram recovered the same major doryctine clades obtained in
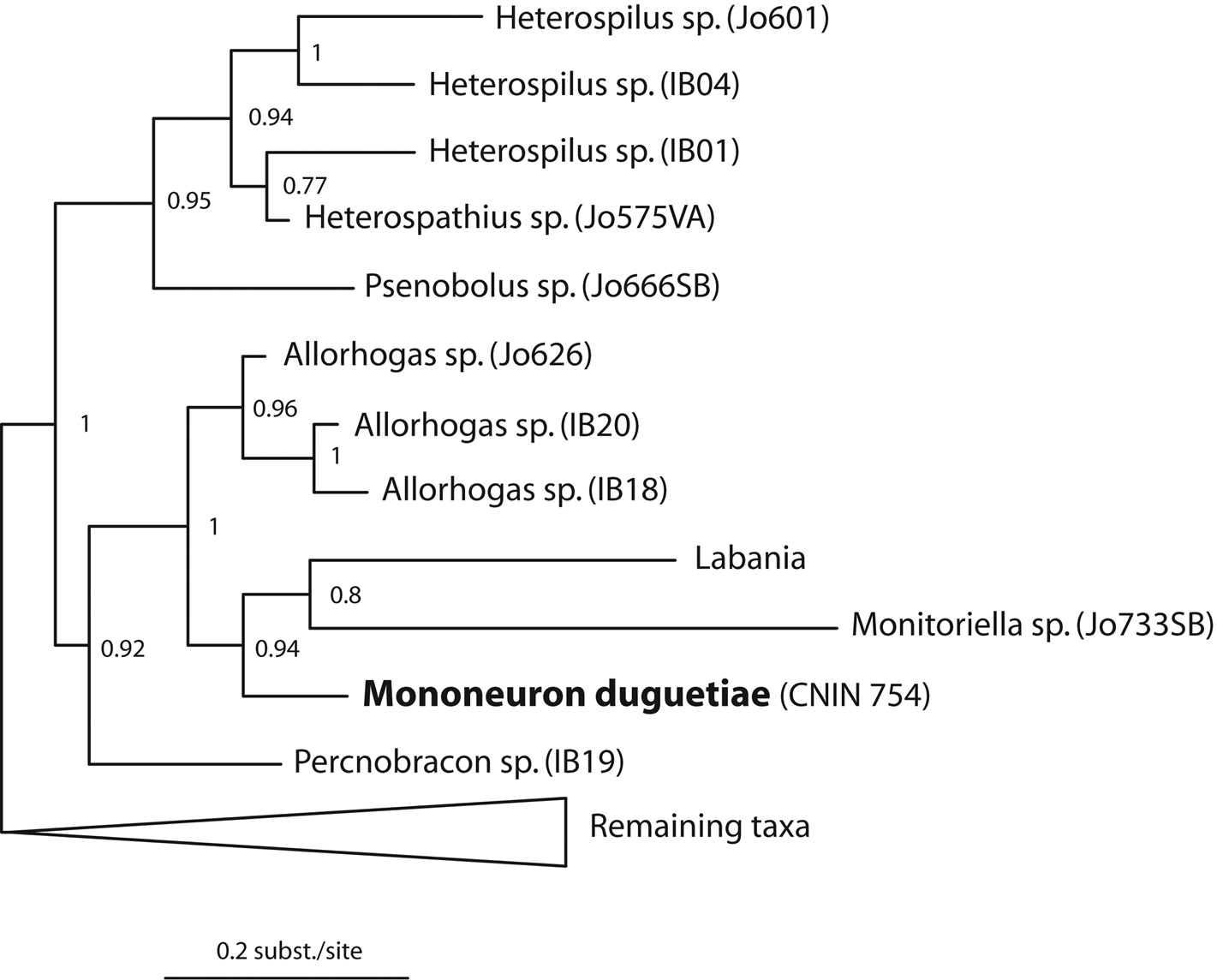
Part of theBayesian phylogram obtained in this study showing the phylogenetic affinities of Mononeuron duguetiae Fischerwith respect to other gall associated doryctine genera (see results for details of the complete reconstructed topology). Numbers near clades represent posterior probabilities.
The Bayesian test showed that the alternative topology with the three included specimens of Allorhogas and Mononeuron duguetiae as monophyletic is present in some of our 95% credible set of trees sampled from Bayesian analysis. We therefore cannot statistically reject the paraphyly of Allorhogas with respect to Mononeuron duguetiae.
Taxonomyhttp://species-id.net/wiki/Mononeuron
Mononeuron duguetiae Fischer
Mononeuron may be distinguished from the remaining doryctine genera by having the following combination of features: frons excavated; occipital carina meeting hypostomal carina; propodeum with semicircular or subtriangular shaped basal median areas (Fig. 2C); fore wing vein r-m present (Fig. 3C); first subdiscal cell of fore wing open at apex (Fig. 3C); hind wing vein SC+R absent (Fig. 3D); hind wing vein cu-a absent or spectral (Fig. 3D); hind wing vein 1A incomplete (Fig. 3D); hind wing vein M+CU equal to or slightly greater than vein 1M (Fig. 3D); hind wing vein m-cu slightly curved toward wing apex or straight to wing margin but never angled toward wing base (Fig. 3D); hind coxa with a distinct baso-ventral tubercle; basal sternal plate of first metasomal tergum less than 0, 25 length of tergum; metasoma sculptured on first three terga (Fig. 2C); ovipositor apex strongly sclerotized and with a single nodus (Fig. 2D).

Mononeuron duguetiae Fischer (holotype, female) A head and mesosoma, lateral view B head and mesoscutum, dorsal view C metasoma, dorsal view D ovipositor.
Mononueuron is morphologically similar to Allorhogas, but it can be distinguished from members of this genus by having the vertex at least partially striate (Fig. 2B) (coriaceous in all described species of Allorhogas); hind wing vein SC+R absent (present in all described species of Allorhogas except Allorhogas ingavera Marsh); hind wing vein cu-a absent or spectral (sclerotized or nebulose in most described species of Allorhogas, only spectral in Allorhogas argentinus Brèthes and Allorhogas shawi Marsh); hind wing vein 1A incomplete (always reaching vein cu-a in other described species of Allorhogas); ovipositor long, about 1.5 times length of metasoma (Fig. 3B) (generally shorter than metasoma in most of the described species of Allorhogas, longer than metasoma in Allorhogas gallicola Gahan).

A Galls on Duguetia furfuracea. B–D Mononeuron duguetiae Fischer (holotype, female). B Habitus, lateral view C fore wing D hind wing.
Female. Colour: Body and first three flagellomeres honey yellow, remaining flagellomeres, fifth tarsomeres and ovipositor sheath light brown (Fig. 3B). Body length: 2.5 mm; ovipositor 2.5 mm. Head: Clypeus smooth, face and frons smooth, frons slightly excavated, vertex slightly striate laterally and anteriorly, smooth posteriorly (Fig. 2B), temple and gena smooth (Fig. 2A); eye 1.45 times higher than wide (lateral view); malar space 0.45 times eye height (lateral view); temple 0.46 times eye width (dorsal view); ocular-ocellar distance 3.3 times diameter of lateral ocellus; length of scape 1.4 times its width (frontal view); antenna with 20 flagellomeres. Mesosoma: Length of mesosoma 1.5 its maximum height; pronotum smooth laterally, pronotal collar short (Fig. 2A); pronotal groove weakly scrobiculate, propleuron smooth; mesoscutal lobes coriaceous; notauli shallow but distinct, obscured before scutellum at middle of mesoscutum in a costate-rugose area (Fig. 2B); scutellar disc slightly coriaceous; median length of scutellar sulcus 0.33 times median length of scutellar disc, with six carinae (Fig. 2B); mesopleuron coriaceous, weakly coriaceous medially (Fig. 2A); subalar groove scrobiculate; precoxal sulcus wide, smooth, 0.5 length of mesopleuron; venter of mesosoma slightly coriaceous; metapleuron strongly rugose; propodeum with a pentagonal areola diverging at the base of propodeum, areolar area rugose, basal median areas coriaceous (Fig. 2C). Wings: Fore wing length 2.75 times its maximum width, length of pterostigma 3.2 times its maximum width, vein r about 0.5 times length of vein 3RSa, vein 3RSb reaching the wing margin as a tubular vein; vein m-cu interstitial to vein 2RS, vein 1cu-a distinctly postfurcal with vein 1M (Fig. 3C); hind wing vein SC+R absent; vein cu-a absent (Fig. 3D); vein m-cu nebulous (Fig. 3D); vein 1A reduced, only present basally; vein M+CU about equal length of vein 1M (Fig. 3D). Legs: Fore tibia with a row of 12 spines; hind femur weakly coriaceous, length 3.86 times its maximum width; hind coxa weakly striate dorsally, coriaceous laterally, with a distinct basal tubercle. Metasoma: Length of first metasomal tergum 0.9 times its apical width, median area slightly coriaceous basally, costate-rugose apically, costate-rugose laterally, with complete longitudinal lateral carinae; dorsope present, acrosternite 0.2 times length of first metasomal median tergite; second metasomal median tergite longitudinally costate (Fig. 2C); third metasomal median tergite costate on basal half, smooth on apical half (Fig. 2C); remaining metasomal median tergites smooth; ovipositor 1.8 times length of metasoma. Ovipositor strongly sclerotized apically, with a single nodus (Fig. 2D).
Male. Slightly smaller than female. Body length 2.0–2.5 mm; hind femur swollen, length 2.35 times its maximum width.
Variation. Females: Antenna with 16–21 flagellomeres; scutellar sulcus with 5–6 carinae.
We collected 20 leaves of Duguetia furfuracea infested with galls at the type locality of Mononeuron duguetiae in February of 2011. All leaves were placed in a plastic bag and maintained at room temperature. Each of the collected leaves had between three to 10 spheroidal galls with a maximum diameter of approximately 1 cm (Fig. 3A). Twenty five specimens of Mononeuron duguetiae emerged from the galls a few days after being collected, and two or more individuals were observed to emerge from each gall. Moreover, two specimens of an unidentified cecydomiid species and the following wasp taxa emerged after most of the specimens of Mononeuron duguetiae had emerged: Torymidae (six specimens belonging to an unidentified species), Eurytomidae (13 specimens of an unidentified species assigned to Sycophila Walker, and eight specimens of an unidentified species), Eulophidae (21 individuals of an unidentified species), and Diapriidae (one unidentified specimen). Members of some of these wasp families are usually parasitoids (Torymidae, Eulophidae), whereas others are generally phytophagous (e.g. Eurytomidae). We could not confirm that Mononeuron duguetiae is the species that induces the above galls, though its early emergence suggests that it might be cecidogenic.
Comments. The genus Mononeuron has been mainly characterised by the absence of hind wing vein cu-a. This feature was employed to distinguish the genus in the key to New World doryctine genera provided by
In our Bayesian analysis, Mononeuron duguetiae was more closely related to Monitoriella and Labania than to the species of Allorhogas, although with marginally non- significant support. This contrasts with our morphological examinations, which show that there are various external morphological features in Mononeuron duguetiae shared with species of Allorhogas. The latter genus is a mainly Neotropical and Nearctic group with 33 described and an undetermined number of undescribed species. Among the features shared by Mononeuron duguetiae and members of Allorhogas are an excavated frons, hind wing vein r-m present, pronotal collar very short or absent, propodeum usually with semicircular or subtriangular shaped basal median areas, basal sternal plate of first metasomal tergum less than 0.25 length of tergum; metasoma sculptured on first three terga, hind coxa with small but distinct basal tubercle, fore wing vein r-m present, first subdiscal cell open at apex, and hind wing vein m-cu usually curved toward wing apex, often straight to wing margin but never angled toward wing base. Further molecular phylogenetic studies employing additional markers and more taxa, especially for the species of Allorhogas, will confirm the relationships of Mononeuron duguetiae with respect to other gall associated doryctine genera.
We also examined various specimens assigned to Mononeuron duguetiae deposited in the DCBU collection that show some morphological differences, and which might represent an undescribed species. These specimens were collected in the type locality of Mononeuron duguetiae and all of them were reared from the same type of galls on Duguetia furfuracea. They differ from our concept of Mononeuron duguetiae by having the vertex, mesopleuron and basal areas of propodeum mostly smooth, and the first metasomal median tergite smooth and with brown colour. Further molecular data will allow us to confirm whether this represents an undescribed species of Mononeuron or intraspecific variation within Mononeuron duguetiae.
Holotype (DCBU).–Female. Brazil, São Carlos, São Paulo, geschlüpft von Gallen von Duguetia furfuracea ST. HILL. (Annonaceae). Other examined material: 31 specimens, 12 males, 19 females. Twelve specimens: Brazil, SP, São Carlos, UFSCar, Cerrado forest, 21°58'7.96"S, 47°53'9.68"W, ex. Duguetia furfuracea galls, 26 January 2011, A. Zaldívar-Riverón and J. Nunes coll. Nineteen specimens: same locality as above, 18 March to11 April 2008, A. M. Penteado-Dias, coll.
We especially thank Juan José Martínez for his helpful comments to the manuscript. We also thank Ma. Cristina Mayorga-Martínez for her assistance in the collection at IB-UNAM and Luciana B. R. Fernandes for taking the SEM photographs. This work was supported by a PhD scholarship given by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes; Brazil) to JFN; a grant given by the Fundação de Amparo à Pesquisa do Estado de São Paulo (INCT dos Hymenoptera Parasitoides da Região Sudeste Brasileira; FAPESP, Programa BIOTA, Brazil) to APD; grants given by the Consejo Nacional de Ciencia y Tecnología [proyectos Ciencia Básica CONACyT no. 511, Programa de Cooperación Bilateral México-Argentina (CONICET), red Temática del Código de Barras de la Vida] and by the Ministerio de Ciencia e Innovación (CGL2010-15786; Spain) to AZR; and by a postdoctoral fellowship given by the Universidad Nacional Autónoma de México (UNAM; DGAPA fellowships) to SFC.