





(C) 2013 Simon van Noort. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The liopterid subfamily Mayrellinae is revised for the Afrotropical region including the description of the following nine new species of Paramblynotus Cameron: Paramblynotus alexandriensis Buffington & van Noort sp. n.; Paramblynotus bayangensis van Noort & Buffington sp. n.; Paramblynotus behara van Noort & Buffington sp. n.; Paramblynotus dzangasangha van Noort & Buffington sp. n.; Paramblynotus matele van Noort & Buffington sp. n.; Paramblynotus parinari Buffington & van Noort sp. n.; Paramblynotus ruvubuensis van Noort & Buffington sp. n.; Paramblynotus seyrigi van Noort & Buffington sp. n.; Paramblynotus zohy van Noort & Buffington sp. n. The genus Paramblynotus is recorded from Madagascar for the first time, with representatives of two species-groups being present on the island: the Paramblynotus yangambicolous species-group and the new Paramblynotus seyrigi species-group, which we erect here to accommodate a single, but highly distinctive new species possessing apomorphic character states. The latter species-group is possibly endemic to Madagascar. We provide identification keys to the species-groups and species occurring in the Afrotropical region. Online dichotomous and interactive Lucid keys are available at http://www.waspweb.org/Cynipoidea/Keys/index.htm
Africa, Afrotropical region, Cynipoidea, Hymenoptera, identification key, Liopteridae, Madagascar, Mayrellinae, systematics
The liopterid subfamily Mayrellinae was established by
Biologically the family is still poorly known with knowledge restricted to a couple of published records of association through rearing: two species of Kiefferiella were reared from buprestid (Acmaeodera pulchella) infested logs (
During visits to European museums to image types of Afrotropical Hymenoptera, the first author discovered a field box in the Paris Museum containing an odd assortment of unidentified wasps collected by André Seyrig in Madagascar during the 1930’s. Among this material were specimens of unusual liopterids representing a new species-group and the first record of Paramblynotus from Madagascar. We describe these in this paper along with further new species of the genus that have recently been collected from Africa during inventory surveys undertaken by the first author and Robert Copeland of ICIPE, and provide keys to the Afrotropical species of Paramblynotus. This revision elevates the current total of Afrotropical liopterid species to 61 with the description of nine new species in this paper.
Specimens were point mounted on black, acid-free card for examination (using a Wild M-5 stereomicroscope with incandescent and fluorescent light sources), photography and long term preservation. Images were acquired using the EntoVision® multiple-focus imaging system. This system comprises a Leica® M16 microscope with a JVC® KY-75U 3-CCD digital video camera attached that fed image data to a notebook computer. The program Cartograph® 5.6.0 was then used to merge an image series (representing typically 10-15 focal planes) into a single in-focus image. Lighting was achieved using techniques summarized in
Morphological terminology follows that of
Identification keys were produced in three formats to facilitate accessibility by a range of end-users (
List of Depositories (Abbreviations [codens] after Ross et al. 1993, except for RMCA which was listed as MRAC)
BMNH Natural History Museum, London. Curator: David Notton
MNHN Natural History Museum, Paris. Curator: Claire Villemant
NMKE National Museums of Kenya, Nairobi. Curator: Martha Gikunga
RCPC Robert Copeland personal collection, Nairobi.
RMCA Royal Museum for Central Africa, Tervuren. Curator: Eliane de Coninck
SAMC Iziko South African Museum, Cape Town. Curator: Simon van Noort
SANC South African National Collection of Insects. Curator: Ros Urban.
USNM National Museum of Natural History, Washington DC. Curator: Matt Buffington.
The Liopteridae are represented in the Afrotropical region by two subfamilies: Oberthuerellinae and Mayrellinae, with the former having been recently revised by
This subfamily includes two genera, Kiefferiella and Paramblynotus, with only the latter genus occurring in the Afrotropical region (
http://species-id.net/wiki/Paramblynotus
http://www.waspweb.org/Cynipoidea/Liopteridae/Mayrellinae/Paramblynotus/index.htm
Medium-sized to very small cynipoids. Very small species look superficially like cynipids, but careful attention to the relative size of the metasomal terga will help separate Paramblynotus from cynipids. Some superficially resemble figitids, especially Thrasorinae (not found in Africa), but can be separated from the latter by having a deeply foveate pronotum and mesoscutum, as well as by the diagnostic liopterid metasomal tergal arrangement with an enlarged sixth abdominal tergum (Figs 8A, 13A). Within Afrotropical Liopteridae, Paramblynotus can be distinguished by lacking any scutellar armament, by the lack of any sort of lobe at the base of the tarsal claws, and by the presence of an axillula (= auricula) on the side of the scutellum (Fig. 28C). Two apomorphic characters were proposed by
Online interactive keys to genera of Afrotropical Liopteridae are available at: http://www.waspweb.org/Cynipoidea/Keys/index.htm . A standard dichotomous key is also available online at this URL as well as in
The genus is represented in all biogeographical regions except for the Western Palaearctic and Australia (
The type female of Paramblynotus yangambicolus was captured on a Drypetes gossweileri (Euphorbiaceae) log in Democratic Republic of Congo (Zaire) (
Paramblynotus species are rare in collections. With the exception of Paramblynotus yangambicolus (6F), Paramblynotus fuscapiculus (14F), Paramblynotus zairiensis (2F), Paramblynotus kekenboschi (2F), Paramblynotus jacksoni (5F, 2M), and Paramblynotus scalptus (2F, 1M) the remaining 19 African species of Paramblynotus treated by
Key to species-groups of Afrotropical Paramblynotus
http://www.waspweb.org/Cynipoidea/Liopteridae/Keys/index.htm
 |
||
| 1 | Mesoscutum smooth, shiny with only remnants of transverse costae. Median mesoscutal impression present, reaching halfway to anterior margin. The two scutellar foveae each with four subcarinae creating a transverse row of 10 longitudinally elongate subfoveae. Latero-ventral margin of pronotum angled where it meets lateral pronotal carina. F1 of antenna shorter than F2 | Paramblynotus seyrigi species-group (Paramblynotus seyrigisp. n.) |
 |
||
| – | Mesoscutum foveate-reticulate, or transversely carinate with foveae set in rows, or with rough discontinuous transverse costae produced into irregularly raised and slightly backward pointing teeth. Median mesoscutal impression, if present, obscured by sculpture. Scutellar foveae usually two in number, or less frequently subdivided into a maximum of 6 subfoveae. Latero-ventral margin of pronotum evenly curved. F1 of antenna equal to or longer than F2 | 2 |
 |
||
| 2 | Pronotal crest gradually raised, medially forming a conspicuous slightly backward pointing ridge-like tooth. Mesoscutum with rough discontinuous transverse costae produced into irregularly raised and slightly backward pointing teeth. Speculum longitudinally costate. Median propodeal area not delimited by lateral propodeal carinae, posteriorly foveate-reticulate | Paramblynotus yangambicolus species-group |
 |
||
| – | Pronotal crest flat. Mesoscutum foveate-reticulate or with continuous transverse carinae with foveae set in rows. Speculum smooth, rarely longitudinally costate. Median propodeal area distinctly delimited by lateral propodeal carinae, posteriorly not foveate-reticulate | Paramblynotus trisetosus species-group. |
We erect this new species-group to accommodate a single species that is likely to be a Madagascan endemic. The island is known for its high degree of endemism (
Diagnosis. Paramblynotus seyrigi has closest affinities with the two Oriental species-groups Paramblynotus ruficollis and Paramblynotus punctulatus of
urn:lsid:zoobank.org:act:CBBFE73D-0460-4F92-A10F-DD409A4A2494
http://species-id.net/wiki/Paramblynotus_seyrigi
http://www.waspweb.org/Cynipoidea/Liopteridae/Mayrellinae/Paramblynotus/Paramblynotus_seyrigi.htm
Figures 1, 2, 3HOLOTYPE. Female: Madagascar, Behara, Museum Paris, I·38, [A.] Seyrig (MNHN). PARATYPES. 13F, 9M: same data as holotype, SAM-HYM-P031800 (MNHN, SAMC, USNM); 6F, 4M: Madagascar, Behara, Museum Paris, XI·38, [A.] Seyrig, SAM-HYM-P044099 (MNHN, SAMC, USNM).
Behara (24°57'S, 46°23'E) is situated in south-eastern Madagascar (in the district of Amboasary Sud, which is part of the Anosy Region) near Tôlanaro (formerly Fort Dauphin), the latter was the first French settlement in Madagascar. There are numerous settlements in Madagascar with the name Behara, but based on an annotated copy of the map in Seyrig’s 1932 paper in combination with the fact that he lived in Tôlanaro for many years, the above locality (which is the largest and most well known settlement named Behara) was deemed to be the most likely (Rousse pers. comm.). André Seyrig collected throughout Madagascar from 1921 to 1944, but his collecting effort was focused down the middle of the southern part of the island from Antanarivo to Tôlanaro and east of Antanarivo to the coast (see map in
Madagascar (currently only known from Behara).
Named after André Seyrig (1897–1945) collector of the type series and prolific collector of many other faunal and floral taxa from Madagascar. Noun in the genitive case.
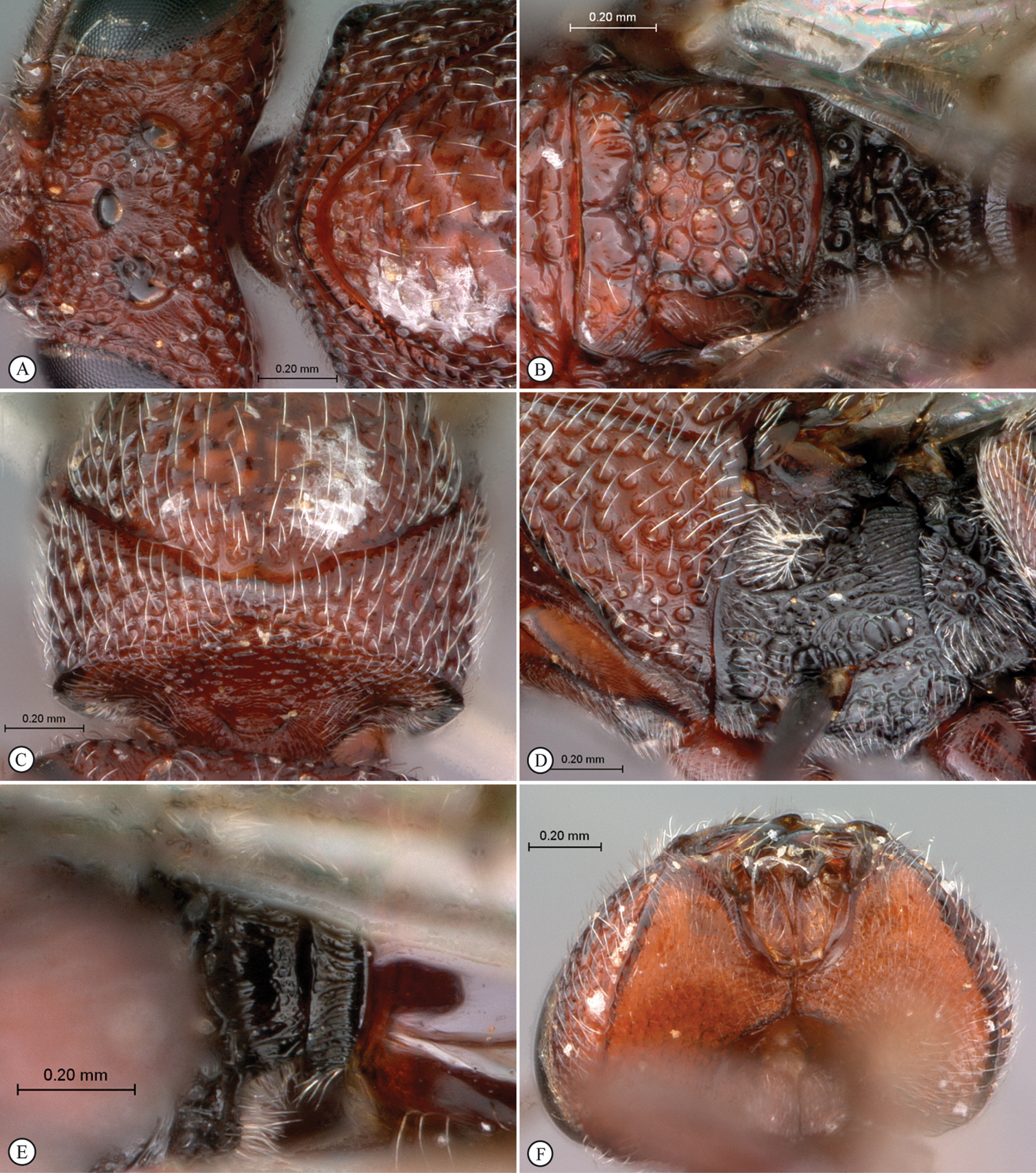
Belongs to the newly erected Paramblynotus seyrigi species-group (see above). Female with 13 (male with 14) segmented antennae (Fig. 1A); F1 shorter than F2; flagellum not widening toward apex; ocellar plate not raised; ocelli large, their diameter as great as distance between lateral and median ocellus. Vertex with two distinct lateral carinae extending from each torulus towards lateral ocelli, reaching posteriorly as far as in line with median ocellus; strongly keeled medial carina present between toruli extending towards median ocellus (Figs 2C; 2E). Occiput straight in dorsal view, smooth without a carina (Figs 1C–D). Lower face protruding in lateral view. Single submedian pronotal depression (Fig. 2A). Pronotal crest not raised. Lateral carina of pronotum distinct. Posterior mesoscutum and scutellum contiguously flat in lateral view (Fig. 1C). Mesoscutum smooth, shiny with remnants of transverse costae; notaulices complete extending to anterior margin of mesocutum; median mesoscutal impression present reaching halfway to anterior margin (Figs 1D–F). The two scutellar foveae each with four subcarinae creating a transverse row of 10 longitudinally elongate subfoveae (Fig. 1F). Upper mesopleuron and speculum glabrous (Fig. 2B). T6 largest dorsally, T8 exposed (Figs 2F; 3A). Wings clear with three darker vertical patches, a small patch either side of the basalis vein, a larger patch subapically between RS and M+Cu and an apical band confluent with the wing margin (Fig. 3B).
This species, along with Paramblynotus zohy sp. n. and Paramblynotus behara sp. n. represent the first records of Paramblynotus from Madagascar.
FEMALE. Length 5.2–8.0 mm (holotype: 6.3 mm ). Head, antennae (except for terminal segments which are darker), pronotum, legs and proximal third of metasoma brownish-orange; rest of mesosoma dark brown; rest of metasoma orange (Fig. 1A). Wings transparent; with three darker vertical patches, a small patch either side of the basalis vein, a larger patch subapically between RS and M+Cu, and an apical band confluent with the wing margin (Fig. 3B). Entire head with the exception of the genae and occiput strongly pubescent. Eyes prominent, bulbous, but not laterally extended much beyond outer margin of genae in frontal view (Fig. 2C). Antenna 13 segmented; F1 shorter than F2; flagellum not widening toward apex. Vertex smooth, ocellar plate not raised; ocelli large, their diameter as great as distance between lateral and median ocellus (Figs 2C; 2E). Face punctate-rugose, humped between toruli and clypeal margin, protruding in lateral view; antennal scrobe mostly smooth with minute punctuation. Vertex with two distinct lateral carinae extending from each torulus, defining outer margin of scrobe, towards lateral ocelli, reaching posteriorly as far as in line with median ocellus; strongly keeled medial carina present between toruli extending towards median ocellus (Figs 2C; 2E). Occiput straight in dorsal view, smooth without a carina. Lower face with strong excavations (with weak vertical carinae) defining upper clypeal margin, and containing anterior tentorial pits (Fig. 2D). Clypeus smooth. Genae with distinct foveae along eye margin, polished between these foveae and genal carina. Mesosoma strongly pubescent. Anterior plate of pronotum puberulous. Pronotum dorsomedially with swollen rim without any crest. Lateral carinae of pronotum strong, fading dorsomedially. Lateral surface of pronotum foveate. Dorsal pronotal area smooth with minute punctures, Mesoscutum smooth, shiny with remnants of transverse costae; notaulices complete, extending to anterior margin of mesocutum; median mesoscutal impression present reaching halfway to anterior margin. The two scutellar foveea each with four subdivisions creating a transverse row of 10 longitudinally elongate subfoveae (Fig. 1F). Scutellum medially smooth with sparse punctures, peripherally areolate-punctate. Posterior mesoscutum and scutellum contiguously flat in lateral view. Mesopleural triangle defined without ventral curved carina, strongly pubescent; upper mesopleuron glabrous, smooth, anteriorly and ventrally pubescent with minute punctures; median longitudinal impression percurrent with evenly spaced transverse carinae; speculum glabrous, smooth (Fig. 2B).
Metanotal-propodeal complex strongly excavated, excavations bordered by strong carinae. Metepisternum dorsally excavated with pubescence, medially polished with indications of minor rugose remnants, ventrally pubescent. Dorsellum with two strong medial foveae; laterally strongly excavated with fine pubescence in lateral depressions. Lateral propodeal carina present; median longitudinal propodeal carina well-defined, crossed by wrinkled transverse and longitudinal carinae extending onto nucha (Fig. 1F). Rs+M of forewing defined, but nebulous where it arises from basal vein at posterior third. Marginal cell 2.8 times as long as wide (Fig. 3B). Abdominal petiole very short, polished, 0.25 times as long as wide in dorsal view (Figs 1C–D). Posterior ventral margin of metasomal T7 sinuate (Fig. 2F). T8 well exposed, with a patch of scattered long setae posteriorly (Fig. 3A). Ventral portions of T2-T7 covered by sternum 3. Relative dorsal length of T3–8: 27:15:15:46:13:8. Tergites dorsally finely punctate; laterally and ventrally polished. All legs sparsely punctuate, strongly pubescent, except metacoxae dorsally glabrous, smooth. Mesotibial outer spur shorter than inner spur; metatibial spurs subequal in length. Ratio of first metatarsomere to the remaining 4 metatarsomeres combined: 0.70.
MALE. Length 4–5.5 mm. Very similar to female except for abdominal petiole, which is much more elongate, T2 as long as wide in dorsal view and twice as long as high in lateral view. Tergites 6–8 each with a dorsal terminal area of setiferous punctures, rest of tergite polished.

Paramblynotus seyrigi sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E mesosoma, dorsal view F scutellum and propodeum, dorsal view.

Paramblynotus seyrigi sp. n., holotype female. A pronotum, antero-dorsal view B mesopleuron C head anterior view D face, anterior view E vertex, dorso-lateral view F metasoma, lateral view.

Paramblynotus seyrigi sp. n., holotype female. A ovipositor lateral view B wings C tarsal claws D labels.
Historically this taxon has been recognized variously at generic (Decellea Benoit, 1956) or subgeneric level (
Previously the species-group was only known from the African mainland with three described species (Paramblynotus mixtus Liu et al., Paramblynotus alveolatus Liu et al. and Paramblynotus yangambicolus Benoit). We describe two further species from Madagascar (Paramblynotus behara sp. n. and Paramblynotus zohy sp. n.).
This species–group is characterized in females by excavations (spiracular peritremata) on the terminal portion of T8 associated with the spiracle (Figs 7B; 9D; 12A–B), not referred to by
Key to species of the Paramblynotus yangambicolus species-group (modified from
| 1 | Forewing with RS+M vein arising at or very close to base of basal vein (Figs 5B; 12C) (Madagascar) | 2 |
| – | Forewing with RS+M vein arising at basal third or mid-way up basal vein (Figs 9B; 9E) (Africa) | 3 |
| 2 | Head and mesosoma black (Figs 4C–D). Abdominal tergite 2 (petiole) dorsally sparsely variolate with confused, weak, longitudinal carinulation (Fig. 4F); laterally with strong, widely spaced longitudinal carinae (Fig. 4E). RS+M, basal vein and M+Cu1 nebulous (Fig. 5B). RS+M arising from base of basal vein, this juncture represented by a pigmented spot (Fig. 5B). (Female unknown) | Paramblynotus behara sp. n. |
| – | Head pronotum, mesoscutum and scutellum reddish-brown (Figs 10 C–D). Abdominal tergite 2 (petiole) densely longitudinally carinate (Fig. 11E). RS+M, basal vein and M+Cu1 distinct (Fig. 12C). RS+M arising slightly above base of basal vein (Fig. 12C). (Female T8 with two excavations, a smaller circular one surrounding the spiracle and a second larger oval one adjacent to the first and extending to posterior margin) (Figs 12A–B) | Paramblynotus zohy sp. n. |
| 3 | Body length about 4 mm. Body colour entirely dark (Fig. 6A). Eyes prominent, protruding distinctly beyond genae (Fig. 6E). Median frontal carina almost reaching clypeus. Antennal scrobes rugose. Speculum distinctly longitudinally carinate (Fig. 6C). Upper mesopleuron foveate-reticulate. Mesoscutellum sloped posteriorly (Fig. 6C). Wings entirely transparent (Fig. 6A). Metasoma somewhat, but not strongly, compressed laterally. T6 almost about the same size as the three preceding tergites (Fig. 6F). Sterna 4-6 entirely covered by sternum 3 | Paramblynotus mixtus |
| – | Body length about 6–10 mm. Head and mesosoma dark, metasoma yellow to brown (Fig. 8A). Eyes not protruding distinctly beyond genae (Fig. 9A). Median frontal carina absent in lower face. Antennal scrobes entirely longitudinally carinate. Speculum very finely and superficially carinate (Fig. 8E). Upper mesopleuron entirely longitudinally costate. Mesoscutellum raised posteriorly, forming a flat dorsal surface (Fig. 8C). Forewing evenly ferruginous with darker marginal cell and a dark narrow strip along anterior-interior margin of the first submarginal cell (Fig. 9B). Metasoma strongly compressed laterally (Fig. 9B). T6 distinctly larger than any of the 3 preceding tergites (Fig. 9C). Sterna 4-6 exposed, not covered by sternum 3 | 4 |
| 4 | Face evenly curved in lateral view. Genae ventrally strongly expanded posteriorly. Median mesopleural impression distinct. Lower mesopleuron densely punctate with pubescence. Apical teeth of metatibia rounded apically; first metatarsomere 2.0× length of remaining 4 metatarsomeres combined | Paramblynotus alveolatus |
| – | Face distinctly raised medially and curved inward ventrally in lateral view (Fig. 8D). Median mesopleural impression usually obscured by extension of longitudinal carinae in upper mesopleuron (Fig. 8C). Lower mesopleuron glabrate and sparsely punctate with sparse pubscence. Apical teeth of metatibia pointed apically (Fig. 9C); first metatarsomere 1.5× length of remaining 4 metatarsomeres combined | Paramblynotus yangambicolus |
Cameroon.
urn:lsid:zoobank.org:act:E8F4B43B-E60E-4A3B-8B58-BCAA65C3A1ED
http://species-id.net/wiki/Paramblynotus_behara
http://www.waspweb.org/Cynipoidea/Liopteridae/Mayrellinae/Paramblynotus/Paramblynotus_behara.htm
Figures 4, 5HOLOTYPE: Male: Madagascar, Behara, Museum Paris, XI·38, A. Seyrig (MNHN). PARATYPES: 1M: same data as holotype, except for I·38 (SAMC); 1M: Madagascar, Bekily, Reg. Sud. de L’ile, Museum Paris, I·39, A. Seyrig (MNHN); 1M: Antsiranana: Orangea, 3km E of Ramena, near fort, 65m, 12°14'49"S, 49°22'17"E, 21–23.I.2001, MT, Irwin, Schlinger & Harin’Hala, littoral forest on sand. MA-01-05-02 (USNM).
Madagascar.
Named after the type locality. Noun in apposition.
Belongs to the Paramblynotus yangambicolus species-group. Male with elongate 14-segmented antenna (Fig. 4A). Forewing with RS+M vein arising at base of basal vein (Fig. 5B), a character state shared with the other Madagascan species in this species-group, Paramblynotus zohy, and separating these two species from the African species. Uniquely, Paramblynotus behara has spectral Rs+M and basal veins (Fig. 5B). Both veins are defined in Paramblynotus zohy; however, this character may be sex-linked and not diagnostic at species level. Males are not known for any of the other species in this species-group.
There is the possibility that this species may be the male of, and hence conspecific, with Paramblynotus zohy sp. n. This is unlikely given the marked sexual dimorphism in overall appearance, a trait that is not characteristic of the genus and one that would need to be invoked if they were conspecific. This species, along with Paramblynotus seyrigi sp. n. and Paramblynotus zohy sp. n., represent the first records of Paramblynotus from Madagascar.
See note under Paramblynotus seyrigi concerning André Seyrig’s collecting.
MALE. Length 2.7 mm. Head and mesosoma black; metasoma reddish-brown; antennae (except for three terminal segments which are darker), are orangish-brown (Fig. 4A). Wings transparent (Fig. 5B). Entire head punctate-rugose, strongly pubescent (Fig. 5A). Eyes small, not laterally extended beyond outer margin of genae in frontal view; 1.17× length of malar space. (Fig. 5A). Antenna 14-segmented; F1 subequal (0.95×) in length to F2; flagellum equally wide throughout length. Vertex areolet-rugose, ocellar plate raised, not delimited by carinae; ocelli small, median ocellus diameter equal to distance between lateral and median ocellus; large areolet positioned laterad of each lateral ocellus (Figs 4D, 5A). Face areolet-rugose; antennal scrobe rugose. Vertex evenly rugose; no carinae present between toruli and lateral ocelli; strongly keeled medial carina present between toruli; extending from median ocellus (originating as v-shaped smooth area at ocellus) to level of ventral margin of toruli (Fig. 5A). Occiput concave in dorsal view, rugose except for smooth, glabrous medial area. Genal carinae extend to mid posterior eye region. Lower face with two weak excavations at upper clypeal margin, containing anterior tentorial pits. Clypeus dorsally rugose, ventrally with horizontal carina, above smooth lighter area bordering straight margin. Genae large.
Mesosoma dorsally with scattered long pubescent. Anterior flange and plate of pronotum uniformly areolete-rugulose and pubescent. Pronotum dorsomedially distinctly raised, in lateral view distinctly higher than anterior margin of mesoscutum (Fig. 4C). Pronotal crest prominent, raised into a sharp lighter medial tooth (Fig. 4C). Lateral pronotal carina distinct, fading well before pronotal crest. Lateral surface of pronotum foveate. Mesoscutum foveate-reticulate with foveae set in irregular rows between transverse costae irregularly raised into strong backward pointing teeth (Figs 4C–D). The two scutellar foveae each subdivided by two very weak sub-lateral longitudinal carinae. Mesoscutellum foveate-reticulate; posteriorly with truncate lamella with a straight edge in dorsal view. Mesopleural triangle ventrally defined by a smoothly curved carina; upper mesopleuron foveate-reticulate; median impression vertically carinate; speculum finely longitudinally carinate (Fig. 4C). Metanotum laterally longitudinally excavated with fine pubescence. Dorsellum with three medial foveae. Propodeum coarsely areolet-rugose, laterally pubescent. Lateral propodeal carinae indistinct and inseparable from the longitudinal carinae; median propodeal area areolate-rugose, with elongate cells posteriorly forming parallel longitudinal carinae.
Rs+M of forewing nebulous, arising from the base of basal vein (Fig. 5B). Marginal cell 2.8 times as long as wide. Abdominal petiole narrow, laterally longitudinally carinate (Fig. 4E), dorsally sparsely variolate with confused, weak, longitudinal carinulation, twice as wide as long in dorsal view (Fig. 4F). Posterior ventral margin of metasomal T6 straight and T7 weakly sinuate in lateral view. T7 largely concealed behind T6 in lateral view, with strong setate medially. T8 strongly setose, visible in lateral view. Relative dorsal length of T3–T8: 7:3.4:3:2:2:3.5. Tergites 5-8 finely punctate; T3-4 polished (Fig. 4F). Legs strongly pubescent; coxae, femora smooth, shiny; tibiae and tarsi densely punctate (Figs 4A, 4E). Mesotibial and metatibial outer spur longer than inner spur. First metatarsomere half as long as the remaining 4 metatarsomeres.
FEMALE. Unknown.

Paramblynotus behara sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E metasoma, lateral view F metasoma, dorsal view.

Paramblynotus behara sp. n., holotype female. A face, anterior view B forewing C labels. Paratype female (Bekily) D habitus, lateral view.
Kenya.
Democratic Republic of Congo, Uganda.

Paramblynotus mixtus, holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E head anterior view F metasoma, lateral view.

Paramblynotus mixtus, holotype female. A antenna, card mount B metasoma, terminal segments C labels; Paramblynotus yangambicolus holotype female. D wings, slide mount.

Paramblynotus yangambicolus, holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E mesosoma, lateral view F mesosoma, dorsal view.

Paramblynotus yangambicolus, holotype female. A head, anterior view B forewing and metasoma, dorsal view C metasoma, lateral view D tergites 6 & 7 and ovipositor, lateral view E wings, slide mount F labels.
urn:lsid:zoobank.org:act:1412C9B3-9C01-454C-A10B-C23C8E586816
http://species-id.net/wiki/Paramblynotus_zohy
http://www.waspweb.org/Cynipoidea/Liopteridae/Mayrellinae/Paramblynotus/Paramblynotus_zohy.htm
Figures 10, 11, 12HOLOTYPE: Female: Madagascar, Bekily, Reg. Sud. de L’ile, Museum Paris, XII·38, A. Seyrig (MNHN).
Distribution: Madagascar (currently only known from Bekily).
Zohy is Malagasy for cave or cavern, with reference to the pothole or cave-like excavations on each side of the terminal portion of T8. Noun in apposition.
Belongs to the Paramblynotus yangambicolus species-group. Female with 13 segmented antennae; F1 same length as F2; flagellum widening towards apex; ocellar plate raised; ocelli small, median ocellus diameter 0.62× distance between lateral and median ocellus (Fig. 11A). Vertex rugose with two weak lateral carinae extending from each torulus towards lateral ocelli; strongly keeled medial carina present between toruli extending from median ocellus and fading below toruli (Fig. 10E). Occiput concave in dorsal view, alveolate without carinae. Upper face protruding in lateral view (Fig. 10C). Pronotal crest distinctly raised into conspicuous medial tooth (Fig. 10C). Lateral carinae of pronotum distinct. Mesoscutum with strongly toothed and ridged transverse costae; notaulices obscured (Figs 10C–D). The two scutellar foveae polished, without divisions (Fig. 11B). Upper mesopleuron and speculum longitudinally striate (Fig. 11D). T6 not much longer than T4 and T5 (Fig. 10A). T8 exposed with two pothole excavations on each side, a smaller circular one surrounding the spiracle and a second larger oval one adjacent to the first and extending to posterior margin (Figs 12A–B). Wings clear; Rs+M arising from near base of basal vein (Fig. 12C), a character state shared with Paramblynotus behara.
This species along with Paramblynotus seyrigi sp. n. and Paramblynotus behara sp. n.represent the first records of Paramblynotus from Madagascar.
See note under Paramblynotus seyrigi concerning André Seyrig’s collecting.
FEMALE. Length 4.5 mm. Head, antennae (except for terminal segments which are darker), mesosoma (except for propodeum and mesopleuron which are black) reddish-brown; legs and metasoma dark brown. Wings clear (Fig. 10A). Entire head punctate-rugose, strongly pubescent. Eyes small, not extending beyond outer margin of genae in anterior view (Fig. 10E); 1.4× length of malar space. Antenna 13 segmented; F1 same length as F2; flagellum widening toward apex. Vertex alveolate, ocellar plate raised; ocelli small, median ocellus diameter 0.62× distance between lateral and median ocellus (Fig. 11A). Face punctate-rugose, humped between toruli and clypeal margin (Fig. 10E), protruding medially in lateral view (Fig. 10C); antennal scrobe rugose. Vertex rugose with two weak lateral carinae extending from each torulus towards lateral ocelli; strongly keeled medial carina present between toruli extending from median ocellus and fading below toruli (Fig. 10E). Occiput concave in dorsal view, alveolate without carinae. Lower face with two weak excavations at upper clypeal margin, containing anterior tentorial pits. Clypeus rugose, margin strongly convex. Genae large, swollen (Fig. 10C).
Mesosoma dorsally with scattered long pubescent (Fig. 10D). Anterior flange of pronotum weakly punctuate/foveate; plate of pronotum medially glabrate and punctate/foveate with pubescence laterally (Fig. 11C). Pronotum dorsomedially distinctly raised, in lateral view distinctly higher than anterior margin of mesoscutum (Fig. 10C). Pronotal crest prominent, raised into a sharp carina running parallel to mesoscutal margin, fronting a horizontal shelf on posterior prontal margin that comprises longitudinally carinate foveae (Fig. 11A). Lateral pronotal carinae distinct, not reaching pronotal crest dorsomedially. Lateral surface of pronotum foveate with smooth areas (Fig. 11D). Mesoscutum foveate-reticulate with foveae set in irregular rows between transverse costae irregularly raised into strong backward pointing teeth (Figs 10C–D). The two scutellar foveae not subdivided by submedian longitudinal carinae (Fig. 11B). Mesoscutellum foveate-reticulate; posteriorly raised and projected into a truncate lamella with a slight emargination in dorsal view (Fig. 11B). Mesopleural triangle ventrally defined by a smoothly curved carina; upper mesopleuron foveate-reticulate; median impression vertically carinate; speculum finely longitudinally carinate (Fig. 11D). Metapectal-propodeal complex coarsely foveate-rugose laterally with dense pubescence. Dorsellum with two medial foveae; laterally excavated with fine pubescence in lateral depressions. Median propodeal area areolate-reticulate (Fig. 11B).
Rs+M of forewing defined, arising from the base of basal vein (Fig. 12C). Marginal cell 2.7 times as long as wide. Abdominal petiole short, longitudinally carinate, 0.22 times as long as wide in dorsal view (Fig. 11E). Posterior ventral margin of metasomal T6 and T7 sinuate (Fig. 10A). T7 largely concealed benath T6 only partially visible laterally and covered on dorso-posterior central margin. T8 marginally exposed with two pothole excavations on each side, a smaller circular one surrounding the spiracle and a second larger oval one adjacent to the first and extending to posterior margin (Figs 12 A–B). Relative dorsal length of T3–8: 21:13:15:12:0:4. Tergites 4-8 finely punctate; T3 polished. All legs sparsely punctuate, strongly pubescent, except metacoxae dorsally glabrous, smooth. Mesotibial outer spur shorter than inner spur; metatibial spurs subequal in length. Ratio of first metatarsomere to the remaining 4 metatarsomeres combined: 0.65.
MALE. Unknown.

Paramblynotus zohy sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E face, anterior view F head and antenna, dorsal view.
Paramblynotus zohy sp. n., holotype female. A vertex and pronotum, dorsal view B scutellum and propodeum, dorsal view C pronotum, antero-dorsal view D mesopleuron E propodeum, dorsal view F head, ventral view.

Paramblynotus zohy sp. n., holotype female. A tergites 5-7 and ovipositor posterior-lateral view B tergites 6-7 and ovipositor posterior view C wings D labels.
Remarks. This is the most species-rich group within the Afrotropical region with 23 previously described species and a further five species added here. The species-group is only known from the African mainland.
Diagnosis. Species in this group are typically smaller than those in other species-groups, and are the easiest to confuse with Figitidae. They are characterized by having a flat pronotal crest (or, pronotal crest absent); the mesoscutum is foveate-reticulate or with continuous transverse carinae with foveae set in rows looking like saw teeth in lateral view; in most species, the speculum is perfectly smooth (gently striate in Paramblynotus vannoorti); and the median propodeal area is distinctly delimited by lateral propodeal carinae, and posteriorly is not foveate-reticulate. Careful attention to the metasomal sclerites will prevent confusing trisetosus-group Paramblynotus with Figitidae.
http://www.waspweb.org/Cynipoidea/Liopteridae/Keys/index.htm
| 1 | Head compressed longitudinally; occiput not concave in dorsal view. Mesoscutum densely foveate, without transverse carinae | Paramblynotus prinslooi |
| – | Head not compressed longitudinally; occiput distinctly concave in dorsal view. Mesoscutum more or less foveate-reticulate, with or without transverse carinae | 2 |
| 2 | Median frontal carina absent. T6 of female metasoma the largest and T8 distinctly exposed. Median propodeal area without a strong transverse carina | 3 |
| – | Median frontal carina present (may only be weakly represented between toruli). T6 of female metasoma not always the largest; if T6 the largest, then T8 is not exposed. Median propodeal area usually with a strong transverse carina. Occasionally variations occur, but never come in combination of features as the above collate | 7 |
| 3 | Forewing entirely clear | 4 |
| – | Forewing at least ferruginous in marginal cell | 5 |
| 4 | Antennae of female with 11 flagellomeres; apical flagellomere less than twice as long as subapical flagellomere. Pronotal crest medially not raised into a triangular process. Metasoma black | Paramblynotus nigricornis |
| – | Antennae of female with 10 flagellomeres; apical flagellomere longer than twice as long as subapical flagellomere. Pronotal crest medially raised into a small but distinct triangular process. Metasoma brown | Paramblynotus claripennis |
| 5 | Antennal scrobes longitudinally carinate in upper part and glabrous in lower part. Upper mesopleuron glabrous, smooth, shiny. Metasoma black | Paramblynotus samiatus |
| – | Antennal scrobes longitudinally carinate entirely. Upper mesopleuron foveate to rugose. Metasoma brown | 6 |
| 6 | Vertex longitudinally carinate laterally. Pronotal crest medially raised into a small, distinct rounded triangular process. Scutellar foveae without submedian carinae | Paramblynotus maculipennis |
| – | Vertex foveate-reticulate entirely without longitudinal carination. Pronotal crest smoothly flat, without triangular process. Scutellar foveae subdivided by distinct submedian carinae | Paramblynotus townesorum |
| 7 | Distance between posterior ocelli at most twice as large as the distance between posterior ocellus and eye. Metasomal T5 of female normal, T6 the largest; T7 exposed, almost entirely covering T8 | 8 |
| – | Distance between posterior ocelli at least three times the distance between posterior ocellus and eye. Metasomal T5 of female dorsally expanded, being the largest (at least so dorsally); T7 is largely or entirely covered by T6; T8 exposed | 22 |
| 8 | Female flagellum distinctly thicker toward apex; median flagellomeres not or slightly constricted toward ends. Antennae with distal flagellomeres 1–3 black, contrasting to the rest, which are yellow-orange | 9 |
| – | Female flagellum not distinctly thicker toward apex; median flagellomeres distinctly constricted toward ends. Antennae yellow or gradually becoming somewhat darker toward apex, but never with contrasting colors between distal and proximal flagellomeres | 13 |
| 9 | Wings with infuscate patch covering submarginal and marginal cells and extending distally slightly beyond vein RS (Fig. 27E) | Paramblynotus ruvubuensis sp. n. |
| – | Wings clear | 10 |
| 10 | Metacoxae ventrally expanded to form a triangular lobular process | Paramblynotus coxatus |
| – | Metacoxae ventrally not expanded to form a triangular lobular process | 11 |
| 11 | Antennae with apical 3 flagellomeres black (Fig. 25A). Antennal scrobes finely punctate and without longitudinal carinae posteriorly | Paramblynotus parinari sp. n. |
| – | Antennae with either the apical or apical 2 flagellomeres dark-brown to black. Antennal scrobes heavily and densely punctate with longitudinal carinae posteriorly | 12 |
| 12 | Female flagellum with terminal two segments usually dark brown; T9 without dense brush of setae | Paramblynotus fuscapiculus |
| – | Female flagellum usually with only terminal segment darkened (occasionally blending into penultimate segment); female T9 with a distinct brush of setae present, engulfing ovipositor (Fig. 13F) | Paramblynotus alexandriensissp. n. |
| 13 | Dorsal surface of head between toruli and posterior margin of lateral ocelli mostly smooth, shiny with lateral carina of antennal scrobe bound on both sides by smooth areas, and sub-confluent with genal carina (Figs 21E–F) | Paramblynotus matele sp. n. |
| – | Dorsal surface of head between toruli and posterior margin of lateral ocelli sculptured or with diagonal carina, except for antennal scrobes, which may be smooth; lateral carina of antennal scrobe interrupted on vertex and not joining genal carina | 14 |
| 14 | Ocellar plate of head not defined by lateral carinae, and without a small, triangular glabrous area beneath anterior ocellus | Paramblynotus trisetosus |
| – | Ocellar plate of head well defined by lateral carinae, with a small, triangular glabrous area beneath anterior ocellus | 15 |
| 15 | Vertex with distinct longitudinal carination | 16 |
| – | Vertex without distinct longitudinal carination | 20 |
| 16 | Median frontal carina almost extending to clypeus. Ocellar plate either with a row of relatively uniform, large foveae along the lateral carinae delimiting the plate or delimited with two parallel carinae | 17 |
| – | Median frontal carina not or slightly extending in lower face. Ocellar plate delimited only by a simple lateral carina | 18 |
| 17 | Ocellar plate with a row of relatively uniform, large foveae along the lateral carinae delimiting the plate | Paramblynotus carinatus |
| – | Ocellar plate delimited only by two simple parallel lateral carinae (Fig. 15B) | Paramblynotus bayangensis sp. n. |
| 18 | Lateral surface of pronotum longitudinally costate in lower part. Lateral propodeal carinae medially strongly curved. Nucha dorsally longitudinally carinate | Paramblynotus kekenboschi |
| – | Lateral surface of pronotum without longitudinal costa in lower part. Lateral propodeal carinae nearly parallel, if strongly curved then nucha dorsally glabrate | 19 |
| 19 | Lateral propodeal carinae medially strongly curved (Fig. 18F) | Paramblynotus dzangasangha sp. n. |
| – | Lateral propodeal carinae nearly parallel | Paramblynotus zairensis |
| 20 | Median frontal carina not distinctly extending in lower face. Head and mesosoma black. Rs and Rs+M veins of forewing distinct and brown in color | Paramblynotus jacksoni |
| – | Median frontal carina distinctly extending to middle of lower face. Head and mesosoma brown to dark brown. Rs and Rs+M veins of forewing reduced and pale in color | 21 |
| 21 | Antennal scrobes not distinctly depressed. Mesoscutum foveate-reticulate. Head, pronotum, mesoscutum, and mesoscutellum dark brown. Antenna yellow. Lateral occipital carinae well developed and crestlike. Median propodeal area glabrate | Paramblynotus cameroonensis |
| – | Antennal scrobes distinctly depressed. Mesoscutum transversely costate with foveae set in between. Head, pronotum, mesonotum, and mesoscutellum reddish brown. Antenna black, except basal two segments yellow. Lateral occipital carinae not crestlike. Median propodeal area areolate-reticulate | Paramblynotus rwandensis |
| 22 | Wings slightly and evenly tinted | Paramblynotus immaculatus |
| – | Wings with large brown to dark brown macula | 23 |
| 23 | Upper mesopleuron foveate-rugose | Paramblynotus minutus |
| – | Upper mesopleuron glabrous | 24 |
| 24 | Vertex diagonally carinate entirely or heavily punctate medially and diagonally carinate laterally | 25 |
| – | Vertex generally glabrate to glabrous with sparse punctures, and sometimes slightly rugose laterally | 26 |
| 25 | Vertex entirely diagonally carinate. Forewing clear basally and distally with a broad medial smoky band across the marginal and submarginal cells (Fig. 30A). Metasomal T7 of female almost covered by T6 (Fig. 30F) | Paramblynotus vannoorti |
| – | Vertex heavily punctate medially and diagonally carinate laterally. Forewing clear in distal two-fifths and ferruginous basally. Metasomal T7 of female distinctly exposed | Paramblynotus scalptus |
| 26 | Lateral propodeal carinae straight and percurrent; medial propodeal area with one straight and percurrent longitudinal carina | Paramblynotus diminutus |
| – | Lateral propodeal carinae discontinuous, medially interrupted by a large fovea; medial propodeal area with more than one non percurrent longitudinal carinae | 27 |
| 27 | Forewing ferruginous only medially, and clear both proximately and distally. Metasomal T7 of female only slightly exposed, only 1:15 as long as T6 as measured medially on lateral sides | Paramblynotus angolensis |
| – | Forewing clear only distally, and ferruginous in basal two-thirds. Metasomal T7 distinctly visible, about 1:2.5as long as T6 as measured along middle of lateral sides | Paramblynotus antistatus |
urn:lsid:zoobank.org:act:337604FD-A3CF-41B0-B520-5DF31141A24B
http://species-id.net/wiki/Paramblynotus_alexandriensis
Figures 13HOLOTYPE. Female: South Africa, [first label] Alexandria, Cape Province, 22.2.1962, ACC. PE 857; [second label] with Curculionid in log of Ptaeroxylon obliquum; [third label] (Hym. Cynipoidea, Mesocynipinae) Paramblynotus Cameron, 1908, sp., det. Michael Soderlund, 1994; [fourth label] red holotype label (SANC). PARATYPES. 3F: Same data as holotype. Deposited in SANC, SAMC, and USNM.
South Africa.
Named after the Alexandria Forest, which now forms part of the Greater Addo Elephant National Park.
Belongs to the Paramblynotus trisetosus clade within the Paramblynotus trisetosus species-group of
FEMALE. Length 2–2.5 mm. Head, mesosoma and metasoma dark brown; antennae and legs light yellow; terminal segment of antennae dark brown (Fig. 13A). Wings transparent (Fig. 13A). Entire head with the exception of the genae and occiput strongly pubescent (Fig. 13E). Eyes prominent, bulbous, but not laterally extended much beyond outer margin of genae in anterior view (Fig. 13E). Antenna 13 segmented; F1 shorter than F2; flagellum not widening toward apex. Vertex foveate, distinct carinae absent; ocellar plate not raised; ocelli large, their diameter as great as distance between lateral and median ocellus (Fig. 13D). Face punctate-rugose, humped between toruli and clypeal margin, protruding in lateral view; antennal scrobe mostly smooth with minute punctuation. Weakly keeled medial carina present between toruli extending towards median ocellus (Fig. 13E). Occiput concave in dorsal view, smooth without a carina. Lower face with strong excavations (with weak vertical carinae) defining upper clypeal margin, and containing anterior tentorial pits (Fig. 13E). Clypeus gently strigate. Genae with distinct foveae along eye margin, punctate-rugose and densely pubescent between these foveae and genal carina (Fig. 13C). Mesosoma strongly pubescent. Single submedian pronotal depression present. Anterior plate of pronotum puberulous. Pronotum dorsomedially with swollen rim without any crest. Lateral carinae of pronotum strong, fading dorsomedially. Lateral surface of pronotum foveate (Fig. 13C). Dorsal pronotal area smooth with minute punctures. Mesoscutum deeply foveate, setose; notaulices complete, extending to anterior margin of mesocutum; median mesoscutal impression absent (Fig. 13D). The two scutellar foveae each with three subcarina creating a transverse row of 8 longitudinally elongate subfoveae. Scutellum medially foveate, sparsely setose, peripherally areolate-punctate (Fig. 13D). Posterior mesoscutum and scutellum contiguously rounded in lateral view. Mesopleural triangle defined without ventral curved carina, strongly pubescent; upper mesopleuron glabrous, smooth, anteriorly and ventrally pubescent with minute punctures; median longitudinal impression percurrent with evenly spaced transverse carinae; speculum glabrous, smooth (Fig. 13C).
Metanotal-propodeal complex strongly excavated, excavations bordered by strong carinae. Metepisternum dorsally excavated with pubescence, medially polished with indications of minor rugose remnants, ventrally pubescent. Dorsellum with two strong medial foveae; laterally strongly excavated with fine pubescence in lateral depressions. Lateral propodeal carina present; median longitudinal propodeal carina well-defined, crossed by wrinkled transverse and longitudinal carinae extending onto nucha. Rs+M of forewing defined, but nebulous where it arises from basal vein at posterior third (Fig. 13A). Marginal cell 2.5 times as long as wide. Abdominal petiole short, longitudinally striate, 0.5 times as long as wide in dorsal view (Fig. 13C). Posterior ventral margin of metasomal T7 gently sinuate (Fig. 13F). T8 well exposed, with a patch of scattered long setae posteriorly (Fig. 13F). Ventral portions of T2-T7 covered by sternum 3. Tergites dorsally finely punctate; laterally and ventrally polished. All legs sparsely punctuate, strongly pubescent, except metacoxae dorsally glabrous, smooth (Fig. 13F). Mesotibial outer spur shorter than inner spur; metatibial spurs subequal in length. Ratio of first metatarsomere to the remaining 4 metatarsomeres combined: 0.67.
MALE. Unknown.

Paramblynotus alexandriensis sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E head, anterior view F metasoma, lateral view.
Angola.
Democratic Republic of Congo.
urn:lsid:zoobank.org:act:248C3CAF-3D2F-4801-A037-1734951AA8ED
http://species-id.net/wiki/Paramblynotus_bayangensis
Figures 14, 15, 16, 17HOLOTYPE. Female: Central African Republic, Prefecture Sangha-Mbaéré, Réserve Spéciale de Forêt Dense de Dzanga-Sangha, 12.7km 326° NW Bayanga, 3°00.27'N, 16°11.55'E, 420m, 12.v.2001, S. van Noort, Sweep, CAR01-S118, Lowland Rainforest, SAM-HYM-P039816 (SAMC). PARATYPES. 1F: Central African Republic, Prefecture Sangha-Mbaéré, Parc National de Dzanga-Ndoki, 38.6km 173° S Lidjombo, 2°21.60'N, 16°03.20'E, 350m, 22.v.2001, S. van Noort, Sweep, CAR01-S240, Lowland rainforest, SAM-HYM-P039849 (SAMC). 1M: Central African Republic, Prefecture Sangha-Mbaéré, Parc National de Dzanga-Ndoki, Mabéa Bai, 21.4km 53° NE Bayanga, 3°02.01'N, 16°24.57'E, 510m, 4.v.2001, S. van Noort, Sweep, CAR01-S27, Lowland Rainforest, marsh clearing, SAM-HYM-P029388 (SAMC); 1F: Central African Republic, Prefecture Sangha-Mbaéré, Parc National de Dzanga-Ndoki, Mabéa Bai, 21.4km 53° NE Bayanga, 3°02.01'N, 16°24.57'E, 510m, 6–7.v.2001, S. van Noort, Malaise trap, CAR01-M56, Lowland Rainforest, marsh clearing, SAM-HYM-P024994 (USNM); 1F : Central African Republic, Prefecture Sangha-Mbaéré, Parc National de Dzanga-Ndoki, Mabéa Bai, 21.4km 53° NE Bayanga, 3°02.01'N, 16°24.57'E, 510m, 6–7.v.2001, S. van Noort, Malaise trap, CAR01-M60, Lowland Rainforest, marsh clearing, SAM-HYM-P024995 (BMNH); 1F : Central African Republic, Prefecture Sangha-Mbaéré, Réserve Spéciale de Forêt Dense de Dzanga-Sangha, 12.7km 326° NW Bayanga, 3°00.27'N, 16°11.55'E, 420m, 11–12.v.2001, S. van Noort, Malaise trap, CAR01-M93, Lowland Rainforest, SAM-HYM-P025016 (SAMC).
Central African Republic.
Named after the village Bayanga.
Belongs to the monophyletic Paramblynotus trisetosus clade, with Paramblynotus coxatus, Paramblynotus fuscapiculus, rwandensis, Paramblynotus trisetosus, Paramblynotus zairensis, Paramblynotus cameroonensis, Paramblynotus kekenboschi, Paramblynotus jacksoni, and Paramblynotus carinatus (
FEMALE (Figs 14A–F, 15A–F): Length 2.2 mm. Head and mesosoma black, metasoma dark brown; coxae, femora dark brown, tibiae and tarsi yellowish-brown (Fig. 14A). Wings clear (Fig. 15E). Antenna 13-segmented; flagellum thicker apically, distal segment longest and widest with three interspersed rows of multiporous plate sensilla (MPS); median flagellomeres constricted proximally and apically; grading from yellow distally to dark brown apically (Fig. 14C). Vertex with smooth area bordering carinae grading to areolet-rugose towards ocelli (Fig. 14D). Eye prominent, distinctly extended laterally beyond outer margin of genae (Fig. 15A). Ocellar plate raised and defined by two parallel lateral weak carinae delimiting the plate, posteriorly foveate, medially smooth (Fig. 15B). Median frontal carina distinct (but need right lighting to see it) in lower face, reaching to epistomal sulcus and bifurcated posteriorly to delimit a glabrous triangular area beneath anterior ocellus (Figs 15A, 15C). Antennal scrobe glabrous, defined by carina laterally. Whole face coarsely areolet-rugose with pubescence. Anterior tentorial pits distinct, situated in shallow depressions. Clypeus diagonally carinate laterally, with an anterior medial smooth excavation, foveate-rugose posteriorly (Fig. 15C). Genae coarsely foveate-rugose (Fig. 14C). Genal carina extending to behind dorso-posterior eye margin. Occiput glabrous. Anterior flange of pronotum glabrate. Anterior plate of pronotum glabrous and sparsely punctate anteriorly. Pronotum dorsomedially not distinctly raised; pronotal crest medially not raised into a process (Fig. 14C). Lateral pronotal carina distinct, almost meeting pronotal crest dorsomedially. Lateral surface of pronotum foveate-reticulate. Mesoscutum distinctly arched dorsally and foveate-reticulate with indistinct transverse costae (Figs 14C–D). Two scutellar foveae not subdivided by carinae; mesoscutellum foveate-reticulate and sloped posteriorly; posterior margin rounded in dorsal view (Fig. 14D). Mesopleural triangle ventrally well defined by smoothly curved carina and with white pubescence (Fig. 14E). Upper mesopleuron, including speculum, glabrous; median longitudinal impression present with unevenly distributed transverse carinae; lower mesopleuron glabrous and pubescent ventrally (Fig. 14E). Metepisternum foveate-reticulate and glabrous above, with an elevated glabrous, smooth area medially, and conspicuously pubescent ventrally (Fig. 14E). Propodeum overall areolate-reticulate; lateral propodeal carina present and distinctly curved medially; median propodeal area glabrate to rugulose; median longitudinal carina present, a transverse carina present anteriorly, and two submedian longitudinal carinae present posterior to transverse carina (Fig. 14F). Rs+M of forewing nebulous, arising from bottom third of basal vein (Fig. 15E). Marginal cell 2.3 times as long as wide. Bulla on Sc+R1 absent. Abdominal petiole 0.5× as long as high in lateral view 0.33× longer than wide in dorsal view, longitudinally carinate (Figs 14C–D). Relative length of T3–7: 14:7:8:28:8; T3–5 glabrous, smooth; T6 finely punctate with a medial row of long white setae; T7 punctate with an anterior row of long white setae; T8 entirely covered by T7 (Fig. 15D). All coxae and femora smooth shiny with pubescence; pro- and meso- tibiae and tarsi finely punctuate with pubescence; meta-tibiae and meta-tarsomeres densely punctate with pubescence (Figs 14A, 14C, 15D). Four apical teeth on metatibia. Proximal metatarsal segment 0.30× length of distal 4 segments.
MALE (Figs 16A–F; 17A–B): Length 1.8mm. Very similar to female, except for longer abdominal petiole, 1.54× as wide as long in dorsal view, as long as high in lateral view (Figs 16C–D). Tergite 5 laterally expanded and by far the largest tergite, latero-medially 1.5× longer than all other tergites combined (Fig. 16D).

Paramblynotus bayangensis sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E mesopleuron F scutellum and propodeum, posterior-dorsal view.

Paramblynotus bayangensis sp. n., holotype female. A head, anterior view B vertex dorso-lateral view C head ventro-lateral view D propodeum and metasoma lateral view E wings F labels.

Paramblynotus bayangensis sp. n., paratype male. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D propodeum and metasoma lateral view E head and mesosoma, dorsal view F head anterior view.

Paramblynotus bayangensis sp. n., paratype male. A wings B labels.
Cameroon.
Democratic Republic of Congo.
Uganda.
South Africa.
Zimbabwe.
urn:lsid:zoobank.org:act:E0A5D4E5-0141-49B9-9D66-C0AD506A8A30
http://species-id.net/wiki/Paramblynotus_dzangasangha
Figures 18, 19, 20HOLOTYPE. Female: Central African Republic, Prefecture Sangha-Mbaéré, Réserve Spéciale de Forêt Dense de Dzanga-Sangha, 12.7km 326° NW Bayanga, 3°00.27'N, 16°11.55'E, 420m, 13.v.2001, S. van Noort, Sweep, CAR01-S162, Lowland Rainforest, SAM-HYM-P039806 (SAMC). PARATYPE. 1M: Central African Republic, Prefecture Sangha-Mbaéré, Réserve Spéciale de Forêt Dense de Dzanga-Sangha, 12.7km 326° NW Bayanga, 3°00.27'N, 16°11.55'E, 420m, 13.v.2001, S. van Noort, Sweep, CAR01-S148, Lowland Rainforest, SAM-HYM-P039807 (SAMC).
Central African Republic.
Named after the Dzanga-Sangha special forest reserve, which forms part of the Dzanga-Ndoki National Park. Noun in apposition.
Belongs to Paramblynotus trisetosus clade of
FEMALE (Figs 18A–F, 19A–C). Length 2.8 mm. Head and mesosoma black; antenna proximally yellow grading to black distally, legs yellow, and metasoma dark brown (Fig. 18A). Forewing with marginal and submarginal cells ferruginous (Fig. 19B). Antennal F1 1.38× longer than F2 (Fig. 18C). Vertex foveate-reticulate and longitudinally carinate, with medial transverse smooth patch adjacent to occiput (Fig. 18D). Eye normal, extending laterally slightly beyond outer margin of genae in anterior view (Fig. 18E). Ocellar plate raised, defined antero-laterally by a carina. Ocelli large, diameter of median ocellus equal to distance between median and lateral ocellus. Face areolet-rugose with scattered white pubescence; antennal scrobe with fine cross striations, glabrous posteriorly with pubescence anteriorly, outside lateral edge defined by a carina. Median frontal carina weakly present between toruli, not extending onto face (Fig. 18E). Anterior tentorial pits distinct situated in slight depressions. Clypeus anteriorly excavated, margin strongly convex, weakly bilobed (Fig. 18E). Genae foveate-reticulate. Genal carina strong, extending to dorso-posterior eye margin. Occiput glabrous, smooth, shiny. Anterior plate of pronotum glabrous, smooth, shiny with two submedian pronotal depressions. Pronotum dorsomedially not distinctly raised; pronotal crest medially raised into a small sharp ridge (Fig. 18C). Lateral pronotal carina distinct, continuous dorsomedially, but not reaching pronotal crest. Lateral surface of pronotum strongly glabrous-foveate (Fig. 18C). Mesoscutum glabrous-foveate (Figs 18C–D). The two scutellar foveae not divided (Fig. 18D). Dorsal surface of mesoscutellum glabrous-foveate; sloping gradually posteriorly (Fig. 18D). Mesopleural triangle ventrally defined by smoothly curved carina; upper mesopleuron glabrous, smooth, shiny; median longitudinal impression present with evenly spaced transverse carinae; speculum glabrous, smooth, shiny (Fig. 18C). Metanotal-propodeal complex areolate-punctate-rugose with metepisternum areolate-punctate in upper part, smooth medially and pubescent ventrally (Fig. 18C). Lateral propodeal carina present, strongly curved medially; median longitudinal propodeal carina present and crossed by two transverse carinae (Fig. 18F). Posterior medial propodel area and nucha glabrous, smooth. Rs+M of forewing absent except for nebulous distal third (Fig. 19B). Marginal cell 1.8 times as long as wide. Bulla on Sc+R1 present. Abdominal petiole 3.5× as wide as long in dorsal view, 2.5× higher than long in lateral view, longitudinally carinate (Figs 18F, 19A). T6 posterior ventral margin sinuate; posterior ventral margin of T7 evenly curved covering T8 (Fig. 19A). Relative length of T3–8: 20:13:15:40:16:0; T7 sparsely finely punctate; T3–6 smooth, shiny; T6 & T7 medially with a row of long white setae (Fig. 19F). All legs smooth, shiny pubescent, yellow contrasting strongly with body (Figs 18A, 19A). Metatibia apically with four small teeth. First metatarsal segment 0.60× remaining four segments.
MALE (Figs 19D–F, 20A–E). Length 2.7 mm. Very similar to female, except for longer abdominal petiole, 2.2× as wide as long in dorsal view, 1.8× higher than long in lateral view (Figs 19F, 20C). Tergite 5 laterally expanded and by far the largest (Fig. 20C).

Paramblynotus dzangasangha sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E head, anterior view F scutellum and propodeum, posterior-dorsal view.

Paramblynotus dzangasangha sp. n., holotype female. A propodeum and metasoma lateral view B wings C labels. Paratype, male. D lateral habitus E dorsal habitus F head and mesosoma, lateral view.

Paramblynotus dzangasangha sp. n., paratype male. A head and mesosoma, dorsal view B head, anterior view C propodeum and metasoma, lateral view D wings E labels. Paramblynotus matele sp. n., holotype female F labels.
South Africa.
South Africa, Zimbabwe.
Namibia
Cameroon
Democratic Republic of Congo
Democratic Republic of Congo, Kenya.
urn:lsid:zoobank.org:act:BEFF5829-CC82-4A1D-BBB9-1170E434DE4A
http://species-id.net/wiki/Paramblynotus_matele
http://www.waspweb.org/Cynipoidea/Liopteridae/Mayrellinae/Paramblynotus/Paramblynotus_matele.htm
Figures 21, 22HOLOTYPE. Female: Central African Republic, Prefecture Sangha-Mbaéré, Parc National de Dzanga-Ndoki, 38.6km 173° S Lidjombo, 2°21.60'N, 16°03.20'E, 350m, 22.v.2001, S. van Noort, Sweep, CAR01-S240, Lowland rainforest, SAM-HYM-P039849 (SAMC). PARATYPES. 1F: Democratic Republic of Congo, Congo Belge: P.N.A., Rwindi, 1000m, 20 au 24.xi.1934, G.F. de Witt: 773; Paramblynotus trisetosus group, det Ronquist, 1994 (RMCA); 1F: Democratic Republic of Congo, Congo Belge: P.N.G., Miss H. De Saeger, Dedegwa, 17-v-1952, H. De Saeger, 3481; Paramblynotus trisetosus group, det Ronquist, 1994 (RMCA).
Central African Republic, Democratic Republic of Congo.
“Matele” is BaAka for tattoo. The BaAka pygmies, who live in the forests of Cameroon, Central African Republic, Congo, Democratic Republic of Congo and Gabon, use sap from the plant Rothmannia whitfieldii to ink tattoos onto their faces. The conspicuous lateral carinae of the antennal scrobes, which join with the genal carina forming an extensive carina running sub-parallel to the edge of the compound eyes, are reminiscent of these tattoo lines. Noun in apposition.
Belongs to the Paramblynotus trisetosus clade within the Paramblynotus trisetosus species-group of
FEMALE. Length 1.9 mm. Head and dorsal mesosoma blackish-brown; lateral mesosoma, metasoma, and coxae dark brown; femora lighter brown, tibiae and tarsi yellowish-brown (Fig. 21A). Wings clear (Fig. 22F). Antenna 13-segmented in paratype (broken in holotype), proximally yellowish-brown gradually darkening towards apex; flagellum slightly thicker apically, distal segment longest and widest with three interspersed rows of multiporous plate sensilla (MPS); median flagellomeres constricted proximally and apically. Vertex posteriorly weakly areolet-rugulose, with weakly defined longitudinal carinae; anteriorly polished (Figs 21E–F). Eye prominent, distinctly extended laterally beyond outer margin of genae (Fig. 22B). Ocellar plate raised, with very weak lateral reticulate carinae; posteriorly weakly areolet-rugose, but largely polished (Figs 21E–F). Median frontal carina weakly defined between toruli extending to just below toruli. Antennal scrobe smooth, polished with isolated setae. Lateral carinae of the antennal scrobes bound by smooth areas (Fig. 21E) and subconfluent with genal carina on the vertex (Fig. 21F). Whole face and genae very weakly areolet-rugulose tending towards being polished, with pubescence (Fig. 22B). Anterior tentorial pits inconspicuous, situated in shallow depressions. Clypeus smooth, with an anterior medial depression (Fig. 22B). Genal carina crested, extending to vertex, where it is subconfluent with the lateral carina of each antennal scrobe (Figs 21E–F). Occiput glabrous, smooth, shiny (Fig. 21D). Anterior plate of pronotum ventro-medially glabrous, polished, laterally and dorsally setose. Pronotum dorsomedially not distinctly raised into a process (Fig. 21C). Lateral pronotal carina distinct, fading dorsomedially. Lateral surface of pronotum dorsally areolet-rugulose, tending towards being polished ventrally. Mesoscutum distinctly arched dorsally and foveate-reticulate with indistinct transverse costae; notauli not evident (Figs 21C–D, 22A). Two smooth, polished scutellar foveae not subdivided by carinae; mesoscutellum areolet-rugulose and sloped posteriorly (Fig. 21D). Mesopleural triangle ventrally well defined by smoothly curved carina and with white pubescence (Fig. 21C). Mesopleuron, including speculum, glabrous, polished; median longitudinal impression present with transverse carinae; lower mesopleural margin bordered with pubescence (Fig. 21C). Metepisternum areolet-rugose, glabrous anterodorsally, conspicuously pubescent ventrally and posterodorsally; median shiny glabrous area present (Fig. 21C). Propodeum areolate-rugose; lateral propodeal carina weakly curved (Fig. 21D). Median propodeal area posteriorly glabrate to rugulose anteriorly; single reticulate transverse carina present anteriorly. Rs+M of forewing nebulous, arising from middle of basal vein (Fig. 22F). Marginal cell 2.9 times as long as wide. Bulla on Sc+R1 absent. Abdominal petiole 0.5× as long as high in lateral view, 2.2× wider than long in dorsal view, longitudinally carinate (Figs 22C–D). Relative length of T3–7: 11:8:9:29:9; T3–5 glabrous, smooth; T6 smooth with a medial row of long white setae; T7 punctate with a medial row of long white setae; T8 covered by T7; basket-like tuft of setae present on the terminal end of T9 (ovipositor sheaths) (Figs 22 C–E). All coxae smooth shiny with lines of pubescence dorsally and medially; femora smooth, shiny, strongly setose; pro- and meso- tibiae and tarsi finely punctuate with pubescence; meta-tibiae and meta-tarsomeres densely punctate with pubescence (Figs 21A, 21C, 22C). Four dorso-apical teeth on metatibia. Proximal metatarsal segment about two-fifths the length of distal 4 segments combined.

Paramblynotus matele sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E head, dorsal view F head, dorsolateral view.

Paramblynotus matele sp. n., holotype female. A mesosoma, dorsolateral view B head, anterior view C metasoma, lateral view D metasoma, dorsal view E metasomal terminal segments F forewing.
South Africa.
Democratic Republic of Congo.
urn:lsid:zoobank.org:act:C38B3330-3F1C-42A5-A7AF-53BD6FEC11DA
http://species-id.net/wiki/Paramblynotus_parinari
http://www.waspweb.org/Cynipoidea/Liopteridae/Mayrellinae/Paramblynotus/Paramblynotus_parinari.htm
Figures 25HOLOTYPE. Female: Uganda, Kibale National Park, Kanyawara, Makerere University Biological Field Station, 1587m, 0°33.408'N, 30°22.603'E, 30vii-5viii.2005, S. van Noort, UG05-M10, Malaise trap, degradedmid-altitude Rainforest, SAM-HYM-P025019 (SAMC). PARATYPES. 2F: one specimen same data as holotype (USNM); second specimen: Kenya, Western Prov., Kakamega Forest, Rondo, 0°14.13'N, 34°51.87'E, MT, 17–31.VII.2006, R. Copeland (NMKE).
Uganda, Kenya.
The rainforest at Kanyawara, the area around MUBFS (Makerere University Biological Field Station) where the holotype was collected, is classified by foresters as Parinari forest, distinguished on photo aspect maps by the large spreading crowns of Parinari excelsa Sabine, a valuable timber tree. Noun in apposition.
Belongs to the Paramblynotus trisetosus clade within the Paramblynotus trisetosus species-group of
FEMALE. Length 2–2.5 mm. Head, mesosoma black; metasoma, coxae and femora (in part) dark brown; antennae and rest of legs light yellow; terminal segment of antennae dark brown (Fig. 25A). Wings transparent (Fig. 25A). Entire head with the exception of the occiput strongly pubescent (Fig. 25E). Eyes prominent, bulbous, laterally extended slightly beyond outer margin of genae in anterior view (Fig. 25E). Antenna 13 segmented; F1 shorter than F2; flagellum slightly widened toward apex (Figs 25A–B). Vertex foveate, carinae absent; ocellar plate distinctly raised, deeply foveate; ocelli large, their diameter as great as distance between lateral and median ocellus (Fig. 25D). Face punctate-rugose, keeled medially between toruli and clypeal margin; protruding in lateral view; antennal scrobe mostly smooth with minute punctuation (Fig. 25E). Occiput concave in dorsal view, smooth without a carina. Lower face deeply foveate, anterior tentorial pits set into shallow, foveate cavities (Fig. 25E). Clypeus entirely foveate. Genae with distinct foveae along eye margin. Mesosoma strongly pubescent (Figs 25C–D). Single submedian pronotal depression absent; lateral foveae present, open. Anterior plate of pronotum puberulous. Pronotum dorsomedially with swollen rim, crest absent (Fig. 25C). Lateral carinae of pronotum strong, fading dorsomedially. Lateral surface of pronotum foveate. Dorsal pronotal area smooth with minute punctures. Mesoscutum deeply foveate, setose; notaulices complete, extending to anterior margin of mesocutum; median mesoscutal impression reduced to small notch on posterior margin of mesoscutum (Fig. 25D). The two scutellar foveae simple, smooth, with a few setae, separated by scutellar ridge; scutellum entirely foveate, evenly setose (Fig. 25D). Posterior mesoscutum and scutellum contiguously rounded in lateral view. Mesopleural triangle defined by ventral curved carina, strongly pubescent; upper mesopleuron glabrous, smooth, anteriorly and ventrally pubescent with distinct punctures; median longitudinal impression well developed, with evenly spaced transverse carinae; speculum glabrous, smooth (Fig. 25C).
Metanotal-propodeal complex strongly excavated, excavations bordered by strong carinae. Metepisternum dorsally excavated with pubescence, medially polished, ventrally pubescent. Dorsellum with two strong medial foveae; laterally strongly excavated with fine pubescence in lateral depressions. Lateral propodeal carina present, curved medially; median longitudinal propodeal carina well-defined, crossed by wrinkled transverse and longitudinal carinae extending onto nucha. Rs+M of forewing defined, but nebulous where it arises from basal vein at posterior third (Fig. 25B). Marginal cell 2.5 times as long as wide. Abdominal petiole short, longitudinally striate, 0.25 times as long as wide in dorsal view (Fig. 25F). Posterior ventral margin of metasomal T6 gently sinuate. T7 with a linear patch of long setae anteriorly, completely covering T8, except for a small protrusion that has a distinct basket-like tuft of setae (Fig. 25F). Ventral portions of T2-T7 covered by sternum 3. Tergites dorsally finely punctate; laterally and ventrally polished. All legs sparsely punctuate, strongly pubescent, except metacoxae dorsally glabrous, smooth (Figs 25A, 25F). Mesotibial outer spur shorter than inner spur; metatibial spurs subequal in length. Ratio of first metatarsomere to the remaining 4 metatarsomeres combined: 0.60.
MALE. Unknown.

Paramblynotus nigricornis, holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E head anterior view F metasoma, lateral view.
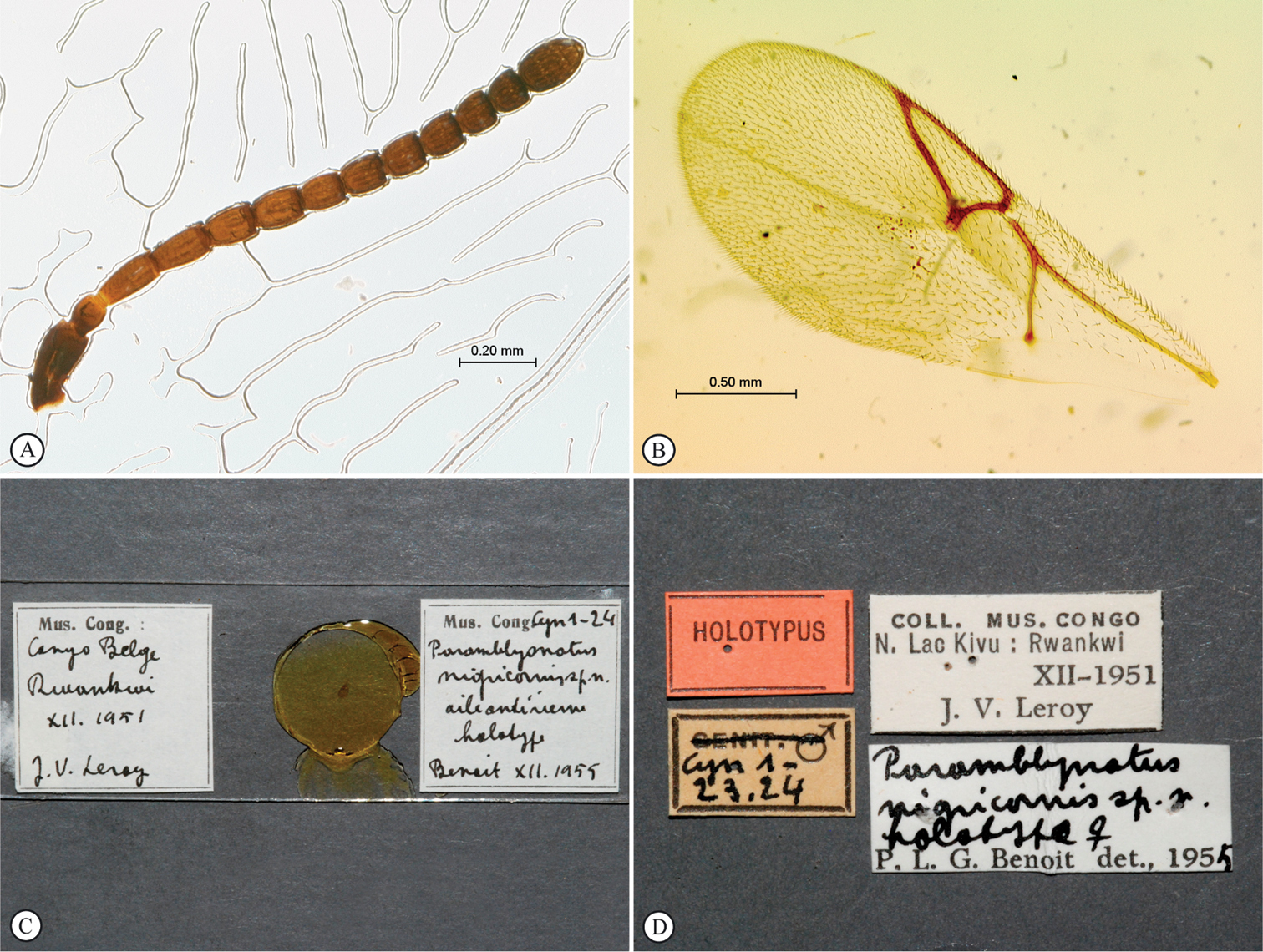
Paramblynotus nigricornis, holotype female. A antenna, slide mount B forewing, slide mount C slide, forewing D labels.

Paramblynotus parinari, holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E head anterior view F metasoma, lateral view.
South Africa.
urn:lsid:zoobank.org:act:8174BAE0-BC7E-4828-8D74-C3B56978DA7F
http://species-id.net/wiki/Paramblynotus_ruvubuensis
Figures 26, 27HOLOTYPE. Female: Burundi, Ruvubu National Park, 1382m, 2.98144°S, 30.45531°E, Malaise trap, edge of forest, near Ruvubu river, 26 Nov–10 Dec 2009, R. Copeland SAM-HYM-P044100 (SAMC).
Burundi.
Named after the type locality along the Ruvubu River in the Ruvubu National Park.
Belongs to the Paramblynotus trisetosus clade within the Paramblynotus trisetosus species-group of
FEMALE. Length 3 mm. Head and mesosoma black, metasoma, coxae, femora dark brown; tibiae and tarsi light brown. Wings clear; weakly infuscate in marginal and submarginal cells extending towards apex slightly past vein RS (Fig. 27E). Antenna 13-segmented; flagellum thicker apically, distal segment longest and widest with three interspersed rows of multiporous plate sensilla (MPS); median flagellomeres constricted proximally and apically; light brown except for last two segments which are blackish-brown (Figs 26A–E). Vertex foveate (Fig. 27C). Eye prominent, distinctly extended laterally beyond outer margin of genae (Fig. 27A). Ocellar plate raised and weakly defined by lateral reticulate carinae; posteriorly foveate, anteriorly areolate (Figs 27A–C). Median frontal carina extending from level of toruli to approximately the level of the ventral eye margins (Fig. 27B). Antennal scrobe glabrous, medially smooth, with fine longitudinal carinae dorsally. Whole face coarsely areolet-rugose with pubescence (Fig. 27A). Anterior tentorial pits distinct, situated in shallow depressions. Clypeus diagonally carinate laterally, with an anterior finely carinate medial excavation (Fig. 27A). Genae coarsely areolet-rugose. Genal carina extending to behind dorso-posterior eye margin. Occiput glabrous, smooth, shiny. Anterior plate of pronotum ventro-medially glabrous, polished, laterally and dorsally setose and sparsely punctate. Pronotum dorsomedially not distinctly raised into a process (Fig. 26C). Lateral pronotal carina distinct, almost meeting pronotal crest dorsomedially. Lateral surface of pronotum areolet-rugulose (Fig. 26C). Mesoscutum distinctly arched dorsally and foveate-reticulate with indistinct transverse costae (Figs 26C–E). Notauli evident posteriorly as smooth depressions with cross carinae (Figs 26D–E). The two scutellar foveae not subdivided by carinae; mesoscutellum areolet-rugose and sloped posteriorly; posterior margin rounded in dorsal view (Fig. 27D). Mesopleural triangle ventrally well defined by smoothly curved carina and with white pubescence. Mesopleuron, including speculum, glabrous, polished; median longitudinal impression present with transverse carinae; lower mesopleural margin bordered with pubescence (Fig. 26C). Metepisternum areolet-rugose and glabrous dorsally, conspicuously pubescent ventrally (Fig. 26C). Propodeum areolate-rugose; lateral propodeal carina curved medially; median propodeal area glabrate to rugulose; median longitudinal carina present, with transverse carina present anteriorly (Fig. 27D). Rs+M of forewing nebulous, arising two-fifths up basal vein (Fig. 27E). Marginal cell 2.2 times as long as wide. Bulla on Sc+R1 absent. Abdominal petiole 0.7× as long as high in lateral view, 1.7× longer than wide in dorsal view, longitudinally carinate (Figs 26C–F). Relative length of T3–7: 14:9:9:18:10; T3–5 glabrous, smooth; T6 finely punctate with a medial row of long white setae; T7 punctate with a medial row of long white setae; T8 mostly covered by T7, but ventro-posteriorly visible (Fig. 26F). All coxae smooth shiny with lines of pubescence dorsally and medially; femora finely punctate, strongly setose; pro- and meso- tibiae and tarsi finely punctuate with pubescence; meta-tibiae and meta-tarsomeres densely punctate with pubescence (Figs 26A, 26C, 26F). Four dorso-apical teeth on metatibia. Proximal metatarsal segment about half the length of distal 4 segments combined.

Paramblynotus ruvubuensis sp. n., holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E hed and mesosoma, dorso-lateral view F metasoma, lateral view.

Paramblynotus ruvubuensis sp. n., holotype female. A head, anterior view B head, antero-lateral view C head, dorsal view D scutellum and propodeum, dorsal view E forewing F labels.
There is a damaged specimen in the Royal Museum for Central Africa (RMCA) collection (Congo Belge: P.N.A., Escarpem. De Kabasha, 1500m, 14.xii.1934, G.F. de Witt: 919; Paramblynotus trisetosus group, det Ronquist, 1994) that possibly may be conspecific with Paramblynotus ruvubuensis, but given the damage we are unable to assign this to a species with any confidence. The metasoma and antennae are missing, as well as the wings and all legs on the right side of the body. Based on interpretation of characters available for observation the specimen appears to be related to Paramblynotus ruvubuensis. The specimen keys to Paramblynotus ruvubuensis, with which it shares the infuscate patch of the forewing.
Rwanda.
South Africa.
South Africa.
Democratic Republic of Congo.

Paramblynotus trisetosus, holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E propodeum and metasoma, lateral view F propodeum and metasoma, dorsal view.

Paramblynotus trisetosus, holotype female. A head, anterior view B forewing, slide mount C antenna, slide mount D slide forewing E slide, antenna F labels.
South Africa.
http://species-id.net/wiki/Paramblynotus_vannoorti
http://www.waspweb.org/Cynipoidea/Liopteridae/Mayrellinae/Paramblynotus/Paramblynotus_vannoorti.htm
Figures 30 A–F1F: Tanzania, Mkomazi Game Reserve, 3°52'59.723"S, 37°52'34.631"E, 30.xii.1995, G. McGavin, Tree canopy fogging, 3/3, Acacia nilotica, SAM-HYM-P025010 (SAMC); 7F, 2M: Tanzania, Mkomazi Game Reserve, 3°52'59.723"S, 37°52'34.631"E, 30.xii.1995, G. McGavin, Tree canopy fogging, 3/2, Acacia reficiens, SAM-HYM-P025011 (SAMC; USNM); 1F: Tanzania, Mkomazi Game Reserve, 3°57'39.399"S, 37°51'44.229"E, 6.i.1996, G. McGavin, Tree canopy fogging, 3/37, Commiphora campestris, SAM-HYM-P025012 (SAMC); 2F: Kenya, Eastern Prov., Samburu Nature Reserve, nr Ewaso Ng’iro River, 874m, 0.56797°N, 37.53563°E, Malaise trap, riverine forest next to headquarters, 18.IX-2.X.2007, R. Copeland (NMKE; USNM); 1F: Kenya, Eastern Prov., Njuki-ini Forest, nr. forest station, 1455m, 0.51660°S, 37.4184°E, Malaise trap just inside indigenous forest, 28.V-11.VI.2007, R. Copeland (RCPC).
Previously only known from the holotype female (Figs 30 A–F), we provide a description of male character states where they differ from those in the female.
MALE. Length 1.5mm. Very similar to female, except for longer abdominal petiole, 1.54× as wide as long in dorsal view, as long as high in lateral view. Tergite 5 laterally expanded and by far the largest tergite, latero-medially 1.5× longer than all other tergites combined. First flagellomere 1.5× length of second, slightly wider distally, excavated laterally.
The Holotype was collected while feeding at flowers of Schotia afra L. (Karoo boer-bean), a tree in the family Fabaceae (subfamily Caesalpinioideae).
Kenya, South Africa, Tanzania.

Paramblynotus vannoorti, holotype female. A lateral habitus B dorsal habitus C head and mesosoma, lateral view D head and mesosoma, dorsal view E head anterior view F metasoma, lateral view.
Democratic Republic of Congo.
The generic richness within the Mayrellinae has oscillated over the years, with the subfamily originally only including the single genus Mayrella. Re-assessment of liopterid subfamily delimitation by
Besides inferred association with Lepidoptera and Coleoptera, and rearing of one species Paramblynotus yangambicolous from a rotten log of Euphorbiacea (Benoit, 1956) (these observations indicating that Paramblynotus species are parasitoids of beetle larva (
The Liopteridae arose between 132 and 90 mya (Buffington et al., 2012). Based on biogeographical dispersal-vicariance analyses in combination with palaeoenvironmental evidence, Liu et al (2007) hypothesized that the African Paramblynotus clade arose as a result of an early dispersal event from the eastern Palaearctic via Arabia during the late Oligocene to early Miocene periods (26–23 mya) with subsequent diversification on the African continent as a result of montane forest fragmentation. Madagascar separated from the African mainland about 160 to 120 mya with India rifting away from Madagascar around 90 mya (
Largely as a result of recent comprehensive targeted inventory surveys (but also as a result of unearthing previously sampled specimens in historical collections) the species richness of the Afrotropical Liopteridae has been elevated from 19 to 61 species with the description of 42 new species over the last 6 years. Since the majority of habitats still remain poorly sampled, we predict that further inventory surveys conducted in the region will reveal additional undescribed species. Implementation of rigorous quantified and replicated surveys across all biomes and vegetation types is a critical necessity in the race to document the region’s biodiversity against a background of rampant habitat destruction and habitat transformation. Further, elucidation of the biology of liopterid wasps of the Afrotropical region is not only critical for understanding the role these wasps play in natural ecosystems, but in agroecosystems as well.
SVN was funded by South African National Research Foundation grants: GUN 2068865; GUN 61497; GUN 79004; GUN 79211; GUN 81139. MLB was funded by the Systematic Entomology Lab, USDA/ARS. The expedition to the Central African Republic was supported by funds from the World Wildlife Fund (US). The Ugandan Wildlife Authority and UNCST provided permits to conduct research in Uganda. We thank Dr. Richard Bagine, Director of Research, Kenya Wildlife Service for granting R. S. Copeland permission to sample in Kenyan National Parks and Reserves. Dr. Helida Oyieke, Head of Collections and Research, National Museums of Kenya, was instrumental in granting permission to loan insects for study. We thank Mattias Forshage (Swedish Museum of Natural History) for his careful review of the manuscript including numerous helpful suggestions and István Miko (Pennsylvania State University) for formulating the URI table. We also acknowledge MorphBank (http://www.morphbank.net ), Florida State University, School of Computational Science, Tallahassee, FL 32306-4026, USA. The USDA is an equal provider and employer.
List of morphological terms. File format: Adobe PDF file (pdf). doi: 10.3897/JHR.31.4072.app
Explanation note: List of morphological terms, their definitions, and corresponding HAO concepts.