





Citation: Ceccarelli FS, Robinson DE, Clebsch H, Zaldívar-Riverón A (2014) Parasitoid wasps from three Jamaican localities: A pilot study. Journal of Hymenoptera Research 37: 127–135. doi: 10.3897/JHR.37.7081
Parasitoid wasps are an extremely speciose, ecologically and economically crucial group of insects. Despite this, they have received disproportionally little attention from scientists, in particular in certain areas of the world. One such area is the Caribbean, where studies are scarce despite the importance of parasitoid wasps, and the uniqueness and diversity of the Caribbean islands. To verify whether an adequate diversity of parasitoid wasps at family level can be found to warrant future studies, this study carries out preliminary sampling in three localities in Jamaica. A total of 1522 individual parasitoid wasps, belonging to at least 16 different families collected during 16 events provide preliminary evidence there is in fact a high diversity of parasitoid wasps in Jamaica, and that future studies there, as in the rest of the Caribbean are definitely worthwhile.
Caribbean, biodiversity, parasitoids
Parasitoids are organisms that spend part of their life cycle feeding on or inside a host, eventually killing it. This lifestyle has been adopted by a notable number of insects, and in Hymenoptera approximately 80% of its species are parasitoids (
The islands of the Caribbean Sea represent a highly interesting part of the world, since they are of complex and varying geological origins: some are volcanic, some tectonic and others continental (
In this study we set out to verify whether collecting efforts in Jamaica would yield high numbers of parasitoid wasps to warrant more detailed future studies, using two of the most common collecting techniques for parasitoid wasps, namely yellow pan trapping (YPT) and sweep netting (e.g.
Parasitoid wasps for this study were collected during the month of November 2010. Three localities were chosen in the Jamaican parishes of St. Andrew, St. Mary and Trelawny (see Figure 1). Within these localities, a total of 16 collecting events took place, either using a sweep net, or setting yellow pan traps (YPT) (for details see Table 1). The three localities, as well as each collecting event within the localities differed in general habitat type, vegetation cover, topology and climatic conditions, all of which were considered when interpreting the results.
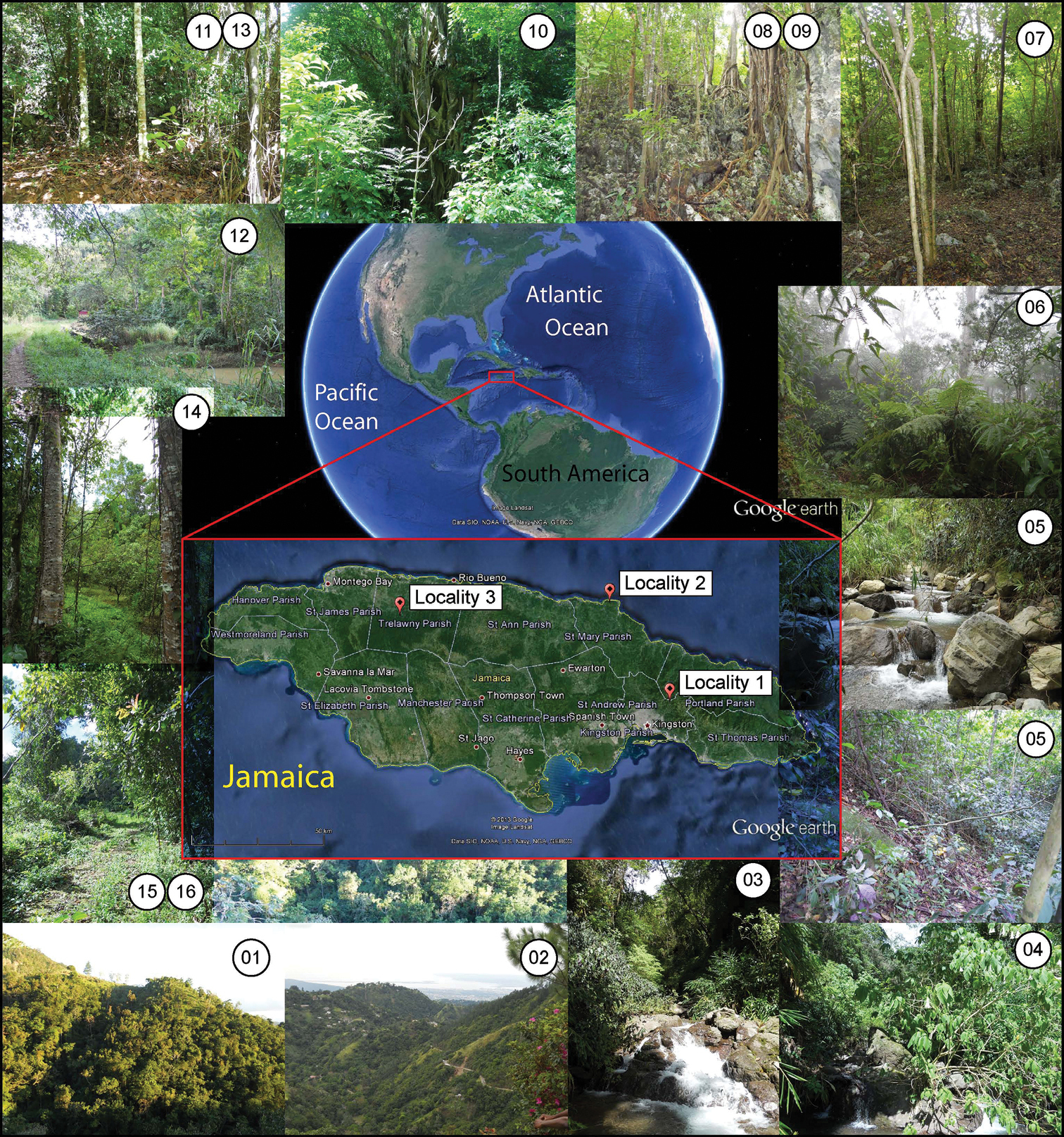
Maps and habitat photographs. Maps in the centre of the figure showing the geographic position of the three localities sampled in this study with surrounding photographs of the habitat types. Numbers in circles represent the collection code as shown in Table 1 without the JAM prefix.
More specifically, locality 1 comprised an area of approximately 0.90 square kilometres (km2) (221.31 acres) in a mountainous area (Blue Mountains), at elevations ranging from 860 to 1205 metres above sea level (m.a.s.l.) As such, most of the collecting sites were on slopes, and the area overall consisted of small patches of primary forest, mostly highly fragmented by human activity. Collecting events JAM01 and JAM03-05 were carried out in the vicinity of fast-flowing mountain streams, with the soil inside the forest patches rich in organic matter. In addition, during collecting events JAM01 to 05 it was sunny, while during event JAM06 there was fog, complicating the sweep netting. Locality 2 consisted of an approximate area of 0.0066 km2 (1.63 acres), at altitudes between 167 and 176 m.a.s.l., with collecting carried out mostly inside woodlands with moderate canopy cover on limestone ground. There was no evident water body in the vicinity. The main disturbing factor during collecting events JAM07 and JAM08 was the heavy rain, which flooded the yellow pan traps left out during the night. Locality 3 consisted of an approximately 0.29 km2 (70.64 acre) area, at altitudes between 82 and 173 m.a.s.l., within Jamaica’s Cockpit Country, an area containing one of the islands’ last contiguous rainforests, although collecting events JAM10-12 and JAM15 took place on the edge of the forest. The only registered water body within the locality was a spring near collecting event JAM15, a tributary to the Martha Brae river. No noteworthy climatic events affected the collecting events JAM10-16.
All parasitoid wasps collected were stored in 96% EtOH and subsequently identified in the laboratory using a dissecting microscope. Identifications were made at least to family level and in some cases to genus. However, due to the lack of expertise for most families and due to time constraints, we are at this moment unable to present data at species (or even genus) level for all families, so in the absence of an equal taxonomic treatment of all families, we present the data at family level. The total number of wasp families was recorded for each locality, and calculations were made adjusting the number of wasp families collected for each site to obtain a unit measurement for collecting effort. For the yellow pan traps the number of wasp families was divided by the number of hours the YPTs were left out, and for the sweep net collection, the number of families was divided by the number of people multiplied by the number of hours spent collecting. All parasitoid wasps collected during this study were deposited at the Colección Nacional de Insectos, Instituto de Biología, Universidad Nacional Autónoma de México, with accession numbers CNIN-JAM0001-1522.
Collection details for this study.
| Code | L | Place | GPS | Alt. | Date | CM |
|---|---|---|---|---|---|---|
| JAM01 | 1 | St. Andrew, Newcastle | 18.06840, -076.71190 | 860 | 09-11.xi.2010 | 50 YPT |
| JAM02 | 1 | St. Andrew, Newcastle | 18.06760, -076.71510 | 956 | 09-11.xi.2010 | 50 YPT |
| JAM03 | 1 | St. Andrew, Newcastle | 18.07127, -076.71813 | 871 | 10.xi.2010 | sweep 11:00–13:00 |
| JAM04 | 1 | St. Andrew, Newcastle | 18.06840, -076.70850 | 1009 | 10.xi.2010 | sweep 15:00–17:00 |
| JAM05 | 1 | St. Andrew, Newcastle | 18.06840, -076.71190 | 860 | 11.xi.2010 | sweep 11:00–12:00 |
| JAM06 | 1 | St. Andrew, Hollywell Park | 18.08609, -076.72629 | 1205 | 11.xi.2010 | sweep 15:30–16:00 |
| JAM07 | 2 | St. Mary, Oracabessa | 18.40207, -076.92519 | 174 | 14-16.xi.2010 | 50 YPT |
| JAM08 | 2 | St. Mary, Oracabessa | 18.40324, -076.92727 | 176 | 15-17.xi.2010 | 50 YPT |
| JAM09 | 2 | St. Mary, Oracabessa | 18.40260, -076.92519 | 167 | 16.xi.2010 | sweep 10:00–11:00 |
| JAM10 | 3 | Trelawny, Windsor | 18.35752, -077.65837 | 97 | 18.xi.2010 | sweep 16:00–17:00 |
| JAM11 | 3 | Trelawny, Windsor | 18.35752, -077.66406 | 82 | 19-21.xi.2010 | 50 YPT |
| JAM12 | 3 | Trelawny, Windsor | 18.35823, -077.65675 | 87 | 19-21.xi.2010 | 50 YPT |
| JAM13 | 3 | Trelawny, Windsor | 18.35169, -077.66371 | 173 | 19.xi.2010 | sweep 14:30–15:30 |
| JAM14 | 3 | Trelawny, Windsor | 18.35531, -077.66371 | 98 | 20.xi.2010 | sweep 11:00–12:00 |
| JAM15 | 3 | Trelawny, Windsor | 18.35838, -077.65837 | 97 | 20.xi.2010 | sweep 15:00–15:30 |
| JAM16 | 3 | Trelawny, Windsor | 18.35838, -077.65837 | 97 | 21.xi.2010 | sweep 15:00–16:30 |
Code = collecting event, L = locality, Place includes Jamaican parish and town, Alt. = altitude in metres within 5 metres, CM = collecting method (YPT = yellow pan traps, sweep = sweep netting, plus the time of day during which sweep netting was carried out).
In this pilot study, we collected a total of 1522 individual parasitoid wasps in the three localities belonging to 15 families and one superfamily (for more details see Table 2 and Suppl. metrial 1). To under- rather than overestimate the number of families, we will count the superfamily, Cynipoidea, as one additional family, making the total number of families collected in this study at least 16. The highest proportion of parasitoid wasps collected in locality 1 belong to the family Diapriidae (especially during the collecting events JAM01 and 02 where we used yellow pan traps), in locality 2 to the family Ichneumonidae and in locality 3 to the Pteromalidae. In total, specimens belonging to 13, 10 and 13 parasitoid wasp families were collected in localities 1, 2 and 3, respectively. Specimens belonging to the families Braconidae, Chalcididae, Cynipoidea (superfamily), Diapriidae, Eulophidae, Ichneumonidae, Platygastridae and Pteromalidae were found in all three localities. Additionally, specimens belonging to the family Mymaridae were only found in localities 1 and 2, specimens belonging to the families Bethylidae and Ceraphronidae were only found in localities 1 and 3, specimens belonging to the family Encyrtidae were only found in localities 2 and 3, specimens belonging to the families Megaspilidae and Proctotrupidae were only found in locality 1 and specimens belonging to the families Agaonidae and Eupelmidae were only found in locality 3.
Number of individual wasps collected from each locality for all families.
| Family | Locality 1 | Locality 2 | Locality 3 | TOTAL |
|---|---|---|---|---|
| Agaonidae | 0 | 0 | 1 | 1 |
| Bethylidae | 2 | 0 | 2 | 4 |
| Braconidae | 196 | 11 | 94 | 301 |
| Ceraphronidae | 1 | 0 | 6 | 7 |
| Chalcididae | 1 | 1 | 5 | 7 |
| Cynipoidea (unidentified families) | 26 | 12 | 91 | 129 |
| Diapriidae | 225 | 10 | 144 | 379 |
| Encyrtidae | 0 | 1 | 3 | 4 |
| Eulophidae | 19 | 1 | 28 | 48 |
| Eupelmidae | 0 | 0 | 1 | 1 |
| Ichneumonidae | 24 | 40 | 8 | 72 |
| Megaspilidae | 1 | 0 | 0 | 1 |
| Mymaridae | 23 | 2 | 0 | 25 |
| Platygastridae | 185 | 4 | 37 | 226 |
| Proctotrupidae | 13 | 0 | 0 | 13 |
| Pteromalidae | 26 | 3 | 275 | 304 |
| TOTAL INDIVIDUALS | 742 | 85 | 695 | 1522 |
| Number of families | 13 | 10 | 13 | 16 |
When accounting for collecting effort, the yellow pan traps were most efficient in locality 1 at family level, meaning that in this locality, using YPTs as a collecting method yielded individuals form more families than in the other two localities. On the other hand, sweep netting was most efficient in locality 3 per unit time (see Figure 2). Parasitoid wasps from most families were collected both in YPTs and sweep nets. The minor differences consisted in the single individuals belonging to the families Agaonidae and Eupelmidae collected by sweep netting but not in YPTs, and the single megaspilid collected by YPT only. Also, in terms of individuals, a higher number of ceraphronids, diapriids, mymarids and platygastrids was collected using YPTs, while for the remaining families more individuals were collected by sweep netting than using YPTs (see Table 3). However, this should not be taken as hard evidence for preferring one method over the other for several reasons. First of all, presenting data in terms of individuals does not equate to species richness; second, collecting effort was not standardised; and thirdly, the numbers we are dealing with in this study are generally low. For these reasons no tests of significance were carried out with regards to differences in productivity between sampling methods.
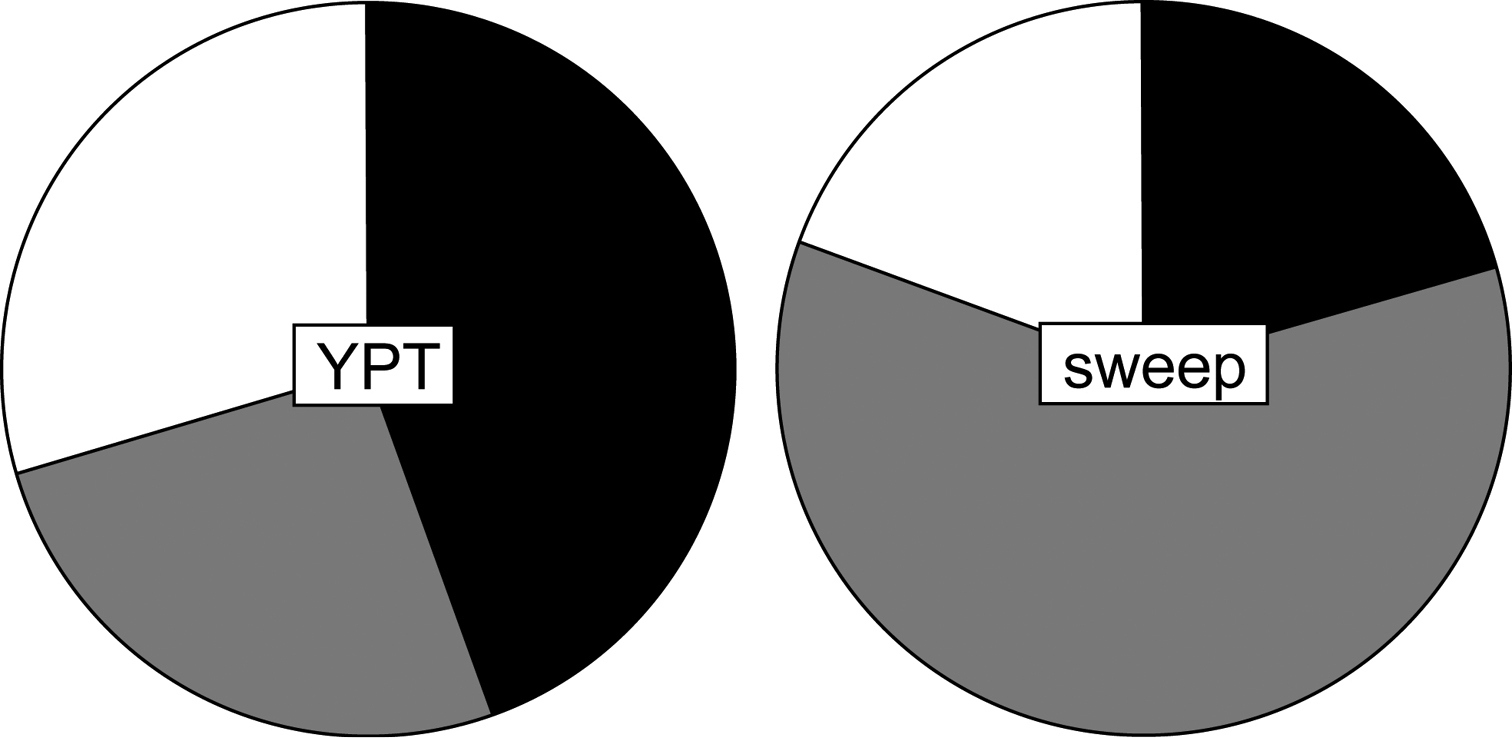
Efficiency of collecting methods. Pie charts representing the efficiency of the two collecting methods used in this study at family level (YPT= yellow pan traps; sweep = sweep net) in units (calculated as described in the Materials and Methods section) for each locality (black= locality 1, grey= locality 2, white= locality 3).
Number of individual wasps collected by the two collecting methods for all families.
| Family | Yellow pan traps | sweep netting |
|---|---|---|
| Agaonidae | 0 | 1 |
| Bethylidae | 1 | 3 |
| Braconidae | 148 | 153 |
| Ceraphronidae | 4 | 3 |
| Chalcididae | 1 | 6 |
| Cynipoidea (unidentified families) | 30 | 99 |
| Diapriidae | 295 | 84 |
| Encyrtidae | 2 | 2 |
| Eulophidae | 22 | 26 |
| Eupelmidae | 0 | 1 |
| Ichneumonidae | 22 | 50 |
| Megaspilidae | 1 | 0 |
| Mymaridae | 23 | 2 |
| Platygastridae | 168 | 58 |
| Proctotrupidae | 7 | 6 |
| Pteromalidae | 151 | 153 |
| TOTAL INDIVIDUALS | 875 | 647 |
| Number of families | 14 | 15 |
The differences between the three localities with regards to the parasitoid wasp families sampled in this study could be attributed to a combination of topology, climate and collecting method. For example, in locality 1 relatively few families were represented in sweep net collections compared to the other two localities. The reasons for this may be that walking the steep slopes and rugged terrain by mountain streams hindered maximum collecting efficiency with a sweep net while the thick fog during collecting event JAM06 definitely lowered the average for the families collected in locality 1. Similarly, the yellow pan traps appeared to be the least efficient in locality 2, however, the heavy rains washing out most of the specimens collected in the traps definitely skewed the results in favour of localities 1 and 3 for YPTs. The fact that the YPTs were most efficient in locality 1 with a high proportion of Diapriidae collected in this locality supports the fact that Diapriidae are efficiently collected using YPTs (e.g.
Given more time and more sampling methods than used in this study, differences in parasitoid wasp family assemblages between localities are more likely to display structuring based on host species assemblages, in turn driven by differences in plant assemblages. Sampling during different seasons throughout the year is also likely to yield different assemblages to those found in this study. Nevertheless, the high numbers of parasitoid wasps belonging to a considerable number of families collected during a relatively short time using only two basic collecting techniques provide evidence that Jamaica is definitely a place worth sampling for future studies on or including these taxa. In addition to collecting for general ecological studies, hymenopterists wishing to undertake studies on specific families may also be able to verify in this pilot study the most appropriate locality for collecting the specific taxon which they wish to investigate.
We thank all the people who facilitated our field work while in Jamaica, especially Oliver Magnus and Time Anson. We also extend our gratitude to Juan José Martínez and Juliano Nunes who helped during the process of wasp identification, and two anonymous referees for comments on a previous version of this manuscript. This work was supported by and a grant given by CONACyT (convocatoria SEP Ciencia Básica 2008) to AZR.
Collection data
Authors: Fadia S. Ceccarelli, Dwight E. Robinson, Hans Clebsch, Alejandro Zaldívar-Riverón
Data type: Collection details
Explanation note: Table showing the number of individual parasitoid wasps collected during this study in each collecting event.
Copyright notice: This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.