





Citation: Popovici OA, Mikó I, Seltmann CK, Deans AR (2014) The maxillo-labial complex of Sparasion (Hymenoptera, Platygastroidea). Journal of Hymenoptera Research 37: 77–111. doi: 10.3897/JHR.37.5206
Hymenopterans have evolved a rich array of morphological diversity within the maxillo-labial complex. Although the character system has been extensively studied and its phylogenetic implications revealed in large hymenopterans, e.g. in Aculeata, it remains comparatively understudied in parasitoid wasps. Reductions of character systems due to the small body size in microhymenoptera make it difficult to establish homology and limits the interoperability of morphological data. We describe here the maxillo-labial complex of an ancestral platygastroid lineage, Sparasion, and provide an ontology-based model of the anatomical concepts related to the maxillo-labial complex (MLC) of Hymenoptera. The possible functions and putative evolutionary relevance of some anatomical structures of the MLC in Sparasion are discussed. Anatomical structures are visualized with Confocal Laser Scanning Microscopy.
Mouthparts, Hymenoptera Anatomy Ontology, Confocal laser scanning microscopy, maxillary palpus, labial palpus, galea, lacinia, prementum, glossa
Despite relatively recent efforts (e.g.,
Due to the extreme diversity of Hymenoptera foraging strategies homologizing mouthpart structures within the order is a challenging task. Other than palpal formulae (
Sparasion is a fairly speciose genus, with 141 valid species (
We examined 34 specimens of 10 Sparasion species (Appendix 1). All specimens were stored in 70% ethanol prior to dissection. Card-mounted voucher specimens are deposited in the Insect Collection of University “Al.I.Cuza” Faculty of Biology, Iasi, Romania (OPPC), in the C. A. Triplehorn Insect Collection, Ohio State University, Columbus, OH, USA (OSUC) and in the Frost Entomological Museum, Pennsylvania State University, State College, PA, USA (PSUC).
We followed the protocols of
MLC specimens were examined under Euromex GE 3045 microscope (400×–1000×). Line drawings were made using Reichart drawing tube attached to the same microscope. Photos were taken using a Leica DFC-500 camera mounted on a Leica M 205A stereomicroscope.
CLSM images were taken on glycerin-stored specimens with Zeiss LSM 710 Confocal Microscope. For visualizing anatomical structures we used excitation wavelength of 488 and emission wavelength of 510-680 nm, detected using two channels visualized separately using two pseudocolors (510–580 nm=green; 580–680 nm=red). For visualizing resilin we used excitation wavelength of 405 nm and emission wavelength of 510–680 nm detected using one channel visualized with blue pseudocolor.
For Scanning Electron Microscopy (SEM) specimens were dried using hexamethyldisilazane (HMDS,
Anatomical concepts used here are defined and aligned with those of
The cardo (cd: Fig. 1A–B) is triangular. The submedial maxillary process of the hypostoma (hys: Fig 1A) inserts submedially on the proximal part of the cardo (caf: Fig. 1B). The mediodistal cardinal ridge, laterodistal cardinal ridge and basal cardinal ridge are present (lcr, mcr, bcr: Fig. 1D) and the inner and outer cardinal processes are absent. The cardo lays almost parallel to the external surface of the hypostoma and is largely obscured by it even if the maxillo-labial complex is fully protracted (Figs 1A, cd: 4A). The conjunctiva connecting the cardo to the rest of the maxillo-labial complex is resilin rich along the stipitocardinal hinge (sch: Fig. 1C).
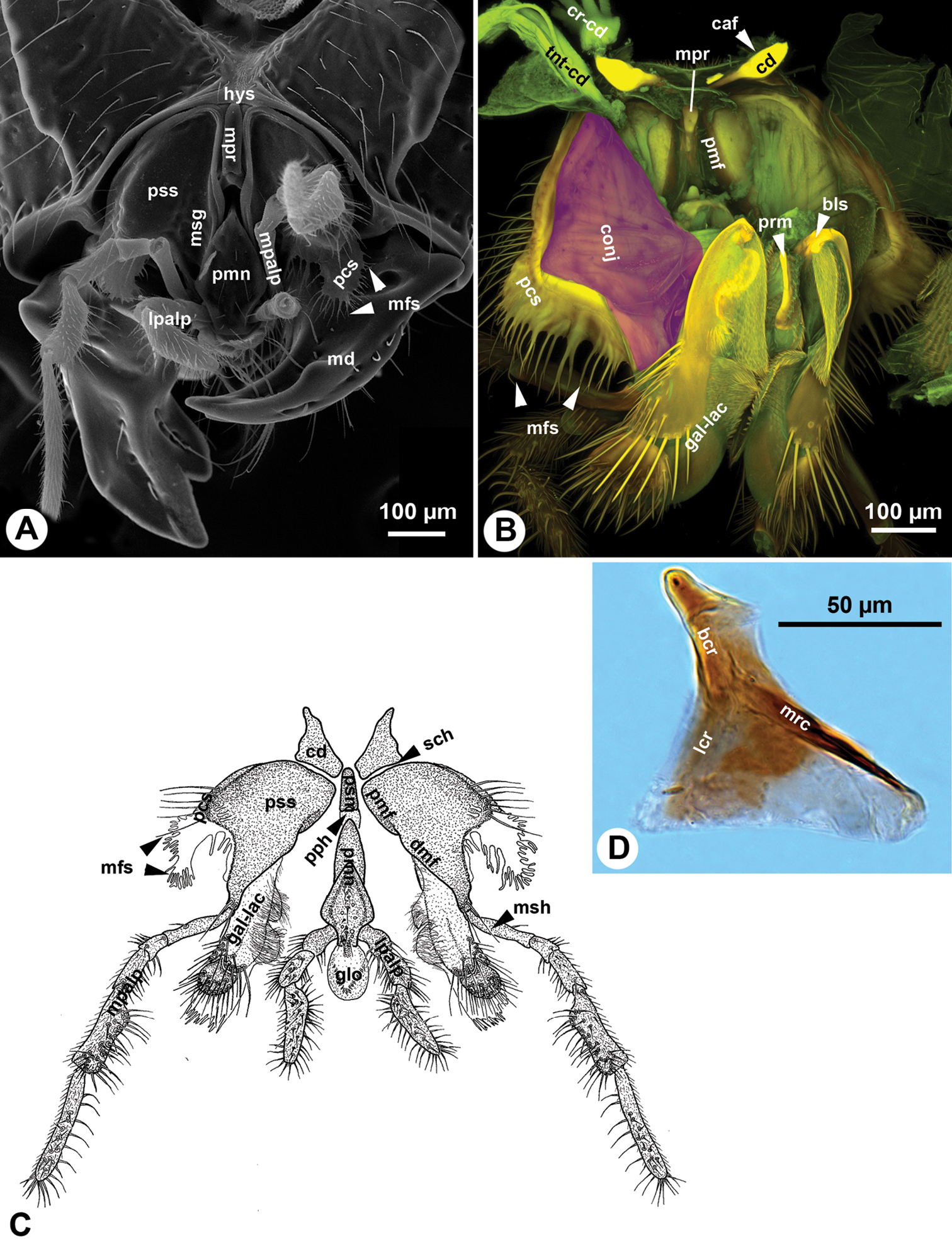
Mouthparts of Sparasion sp. A SEM micrograph showing the mouthparts, posterior (external) view, distal to the bottom B CLSM volume rendered image showing the maxillo-labial complex, anterior (internal) view, distal to the bottom (doi: 10.6084/m9.figshare.861065, doi: 10.6084/m9.figshare.861058) C Line drawing showing the maxillo-labial complex, posterior (external), distal to the bottom D Bright field image showing the cardo of Sparasion, lateral to the left.
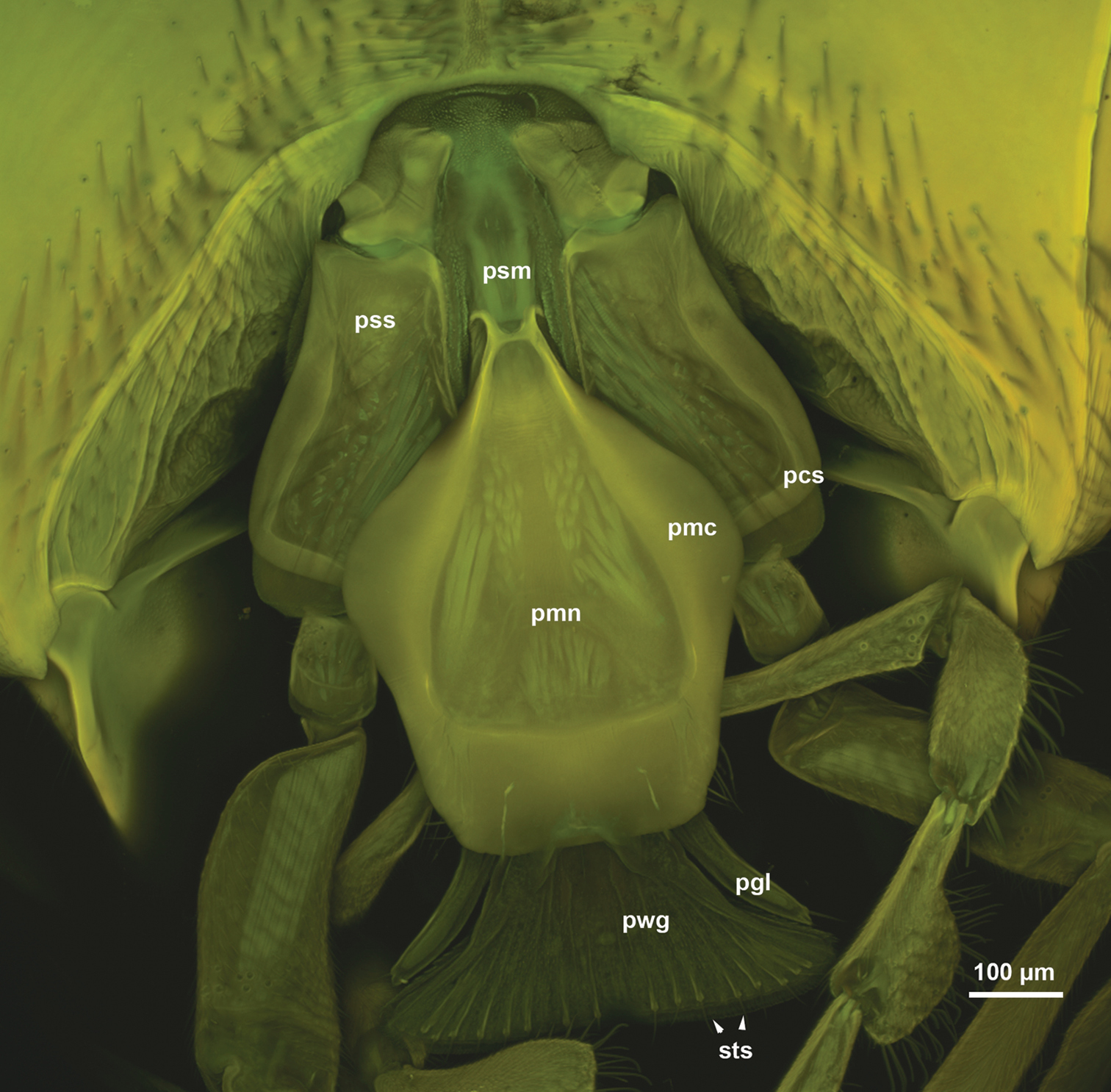
The stipes is triangular in cross section distally. The posterior stipital wall of the stipes bears the posterior stipital sclerite (pss: Figs 1A, C, 2B, C), while the partly sclerotised medial wall and the convex and membranous anterolateral wall (conj: Fig. 1B) bears the galeo-lacinial complex (Fig. 2A, B, D; gal-lac: Fig. 1B, C). The posterior stipital sclerite is triangular in posterior view, and is margined by the principal carina of stipes (pcs: Figs 1A–C, 3A), which is less rigid and melanized than other regions of the sclerite and is the most developed medially and distolaterally. The principal carina of stipes is equipped distolaterally with the marginal fringe of the stipes (mfs: Figs 1A–C, 2B, C) composed of occasionally branched spines. The extent of the principal carina of the stipes on the distolateral margin is variable in different Sparasion species (compare pcs of Fig. 1A with that of Fig. 1B) but it always overlaps the proximomedial surface of the mandible (md: Fig. 1A). The median part of principal carina of the stipes is divided into the proximomedian stipital flange (pmf: Figs 1B, C, 2A) and the distomedial stipital flange (dmf: Figs 1C, 2A, B, C). The proximal part of the distomedial stipital flange posteriorly overlaps the distal part of the proximomedial stipital flange (pmf, dmf: Fig. 1C). The stipes articulates with the postmentum (psm: Figs 1C, 2C) and the distal prementum (pmn: Fig. 1A, C) along the proximomedial stipital flange and with the proximal prementum via the distomedial stipital flange. The medial stipital groove (msg: Figs 1A, 2B) extends medially along the proximomedial stipital flange and distomedial stipital flange and accommodates the first sclerite of the maxillary palp (mpalp: Fig. 1A, C, 3D, E) when it is adpressed against the stipes. The posterior stipital sclerite is glabrous, except a for few, elongate, mechanosensory hairs (msh: Figs 1C, 2B) just proximal of the base of maxillary palp and along the distolateral margin of the stipes abutting the hypostoma. The distal part of the posterior stipital sclerite is equipped with numerous campaniform sensilla (cps: Fig. 2C), which are visible only with transmitted light.
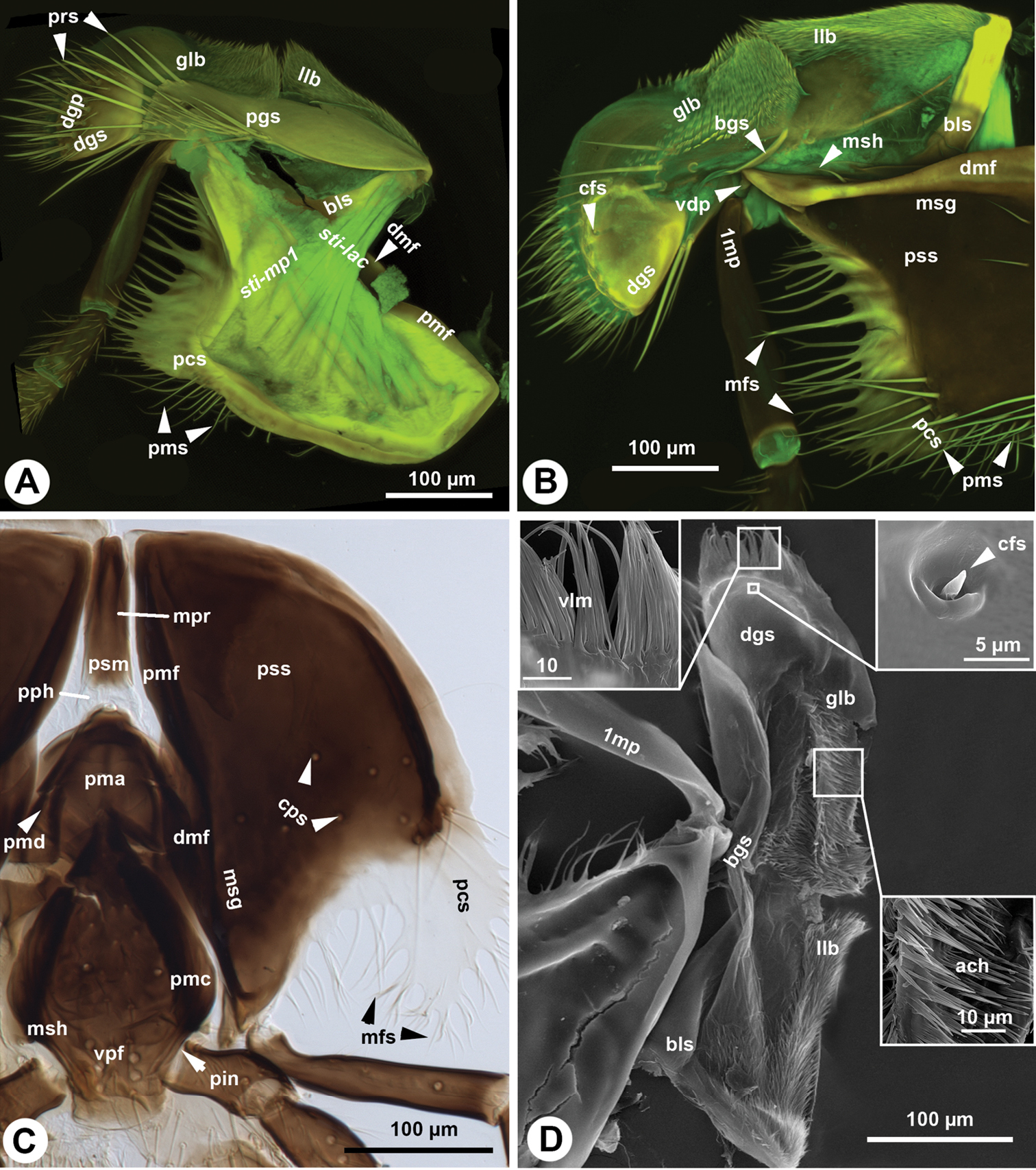
Maxillo-labial complex of Sparasion. A, B CLSM volume rendered images showing the maxilla: A. anterolateral (internal lateral) view (doi: 10.6084/m9.figshare.861060), distal to the left B posteromedial (external medial) view (doi: 10.6084/m9.figshare.861057), distal to the left C Brightfield image showing the maxillo-labial complex, posterior (external) view, distal to the bottom D SEM micrograph showing the maxilla, posteromedial (external-medial) view, distal to the top.
Maxilla and labium of Sparasion. A SEM micrograph showing the skeletomuscular system of the maxilla, posterior (internal) view, distal to the top B SEM micrograph showing the maxillary palp, distal to the top C Bright field image showing the labium, lateral view, distal to the left D, E Bright field image showing the maxillary palps, distal to the left.
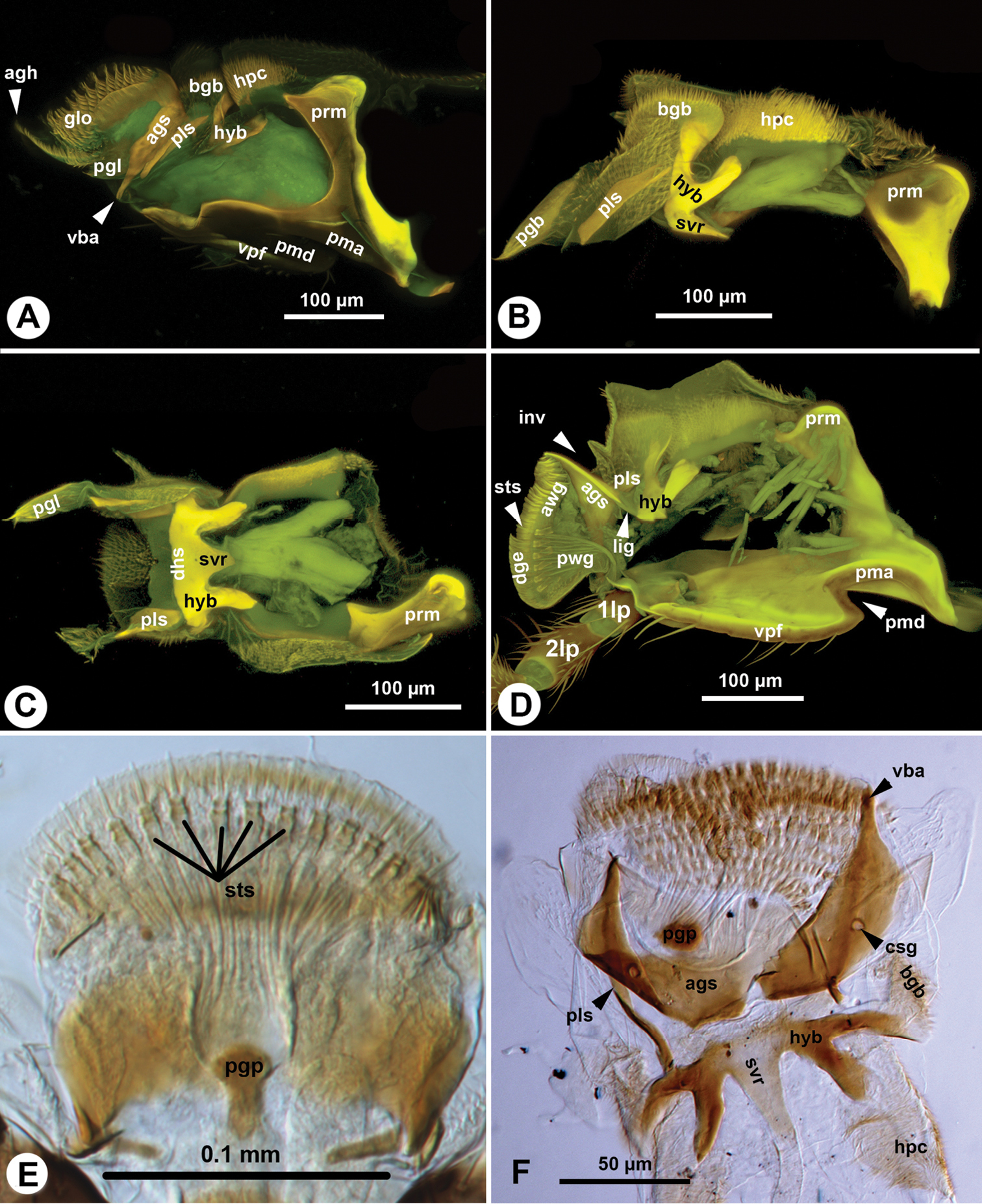
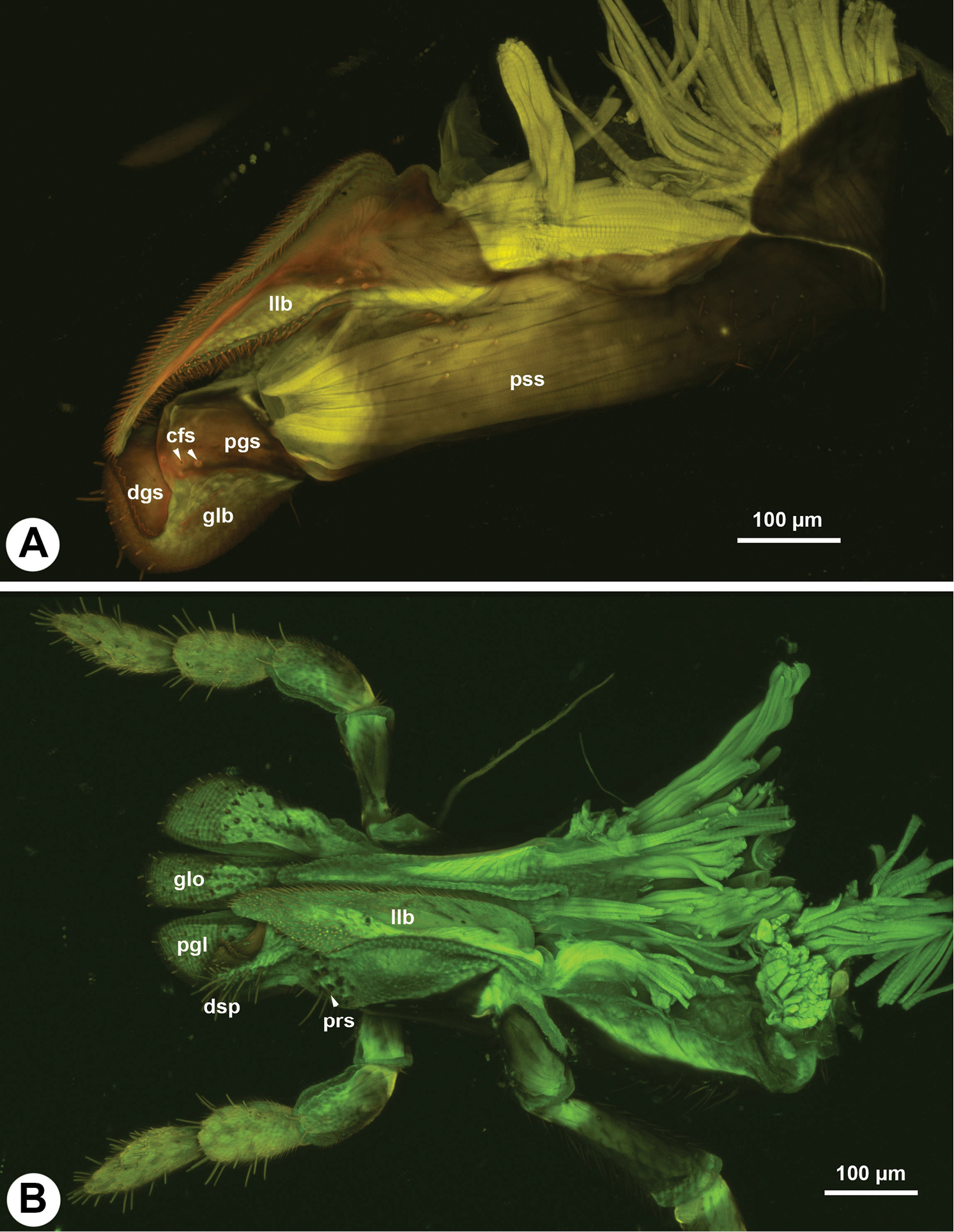
The galeo-lacinial complex has four sclerites and two marginal lobes. The proximal, inverted T-shaped lacinial lever (bls: Fig. 2A, B) and the distal, narrow basal galeal sclerite (bgs: Fig. 2B, D) are situated on the median wall of the stipes and articulate with the posterior stipital sclerite along the distomedial stipital flange. The lacinial bar and lacinial comb are absent. The proximolateral galeal sclerite (pgs: Fig. 2A) is situated in the middle on the lateral wall of stipes and is connected proximally with the lacinial lever and distally with the basal galeal sclerite. The number of mechanosensory hairs in the proximolateral galeal setiferous patch (prs: Fig. 2A) is variable in different Sparasion species (Table 1) and overlaps the distolateral galeal sclerite (dgs: Fig. 2A, B, D), which traverses the galeo-lacinial complex and is represented on both its medial and lateral walls (the complex is unilayered at the sclerite). The proximolateral and distolateral galeal sclerites are connected to each other by the lateral galeal crease. The apicomedial galeal plate is absent. The number of setae in the distolateral galeal setiferous patch (dgp: Fig. 2A) is variable in different Sparasion species (Table 1). The proximal galeal brush is absent. The single coeloconic sensillum of galea (cfs: Fig. 2B, D) is located distally on the median surface of the distolateral galeal sclerite. The single lacinial lobe (llb: Fig. 2A, B, D) extends anteroproximal whereas the galeal lobe (glb: Fig. 2A, B, D) distal to the proximal galeal sclerites. The lacinial lobe and the proximal part of the galeal lobe are covered with short acanthae (ach: Fig. 2D), which comprise the spiculate patch of the lacinia and the spiculate patch of galea respectively. The galeal comb, galeal lamina and galeal fringe are absent. The velum (vlm: Fig. 2D) is fringed distally in some species (vlm: Fig. 2D). The stipital sclerite is absent from Sparasion.
| Characters | Sparasion | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| sp.1 | sp.2 | sp.3 | sp.4 | sp.5 | sp.6 | sp.7 | sp.8 | sp.9 | |
| The number of setae in the proximal galeal brush | 5–7 | 5–7 | 5–7 | 5–7 | 5–7 | 5–7 | 7–10 | 5–7 | 6 |
| The number of setae in the distal galeal setiferous patch | 18–19 | 18–20 | 18–20 | 11–14 | 18–20 | 17 | 27–34 | 18–20 | 22 |
| The number of styloconic sensilla on the glossa | 14–15 | 15–16 | 10–12 | 10–12 | 17 | 23 | 18–19 | 15–16 | 10 |
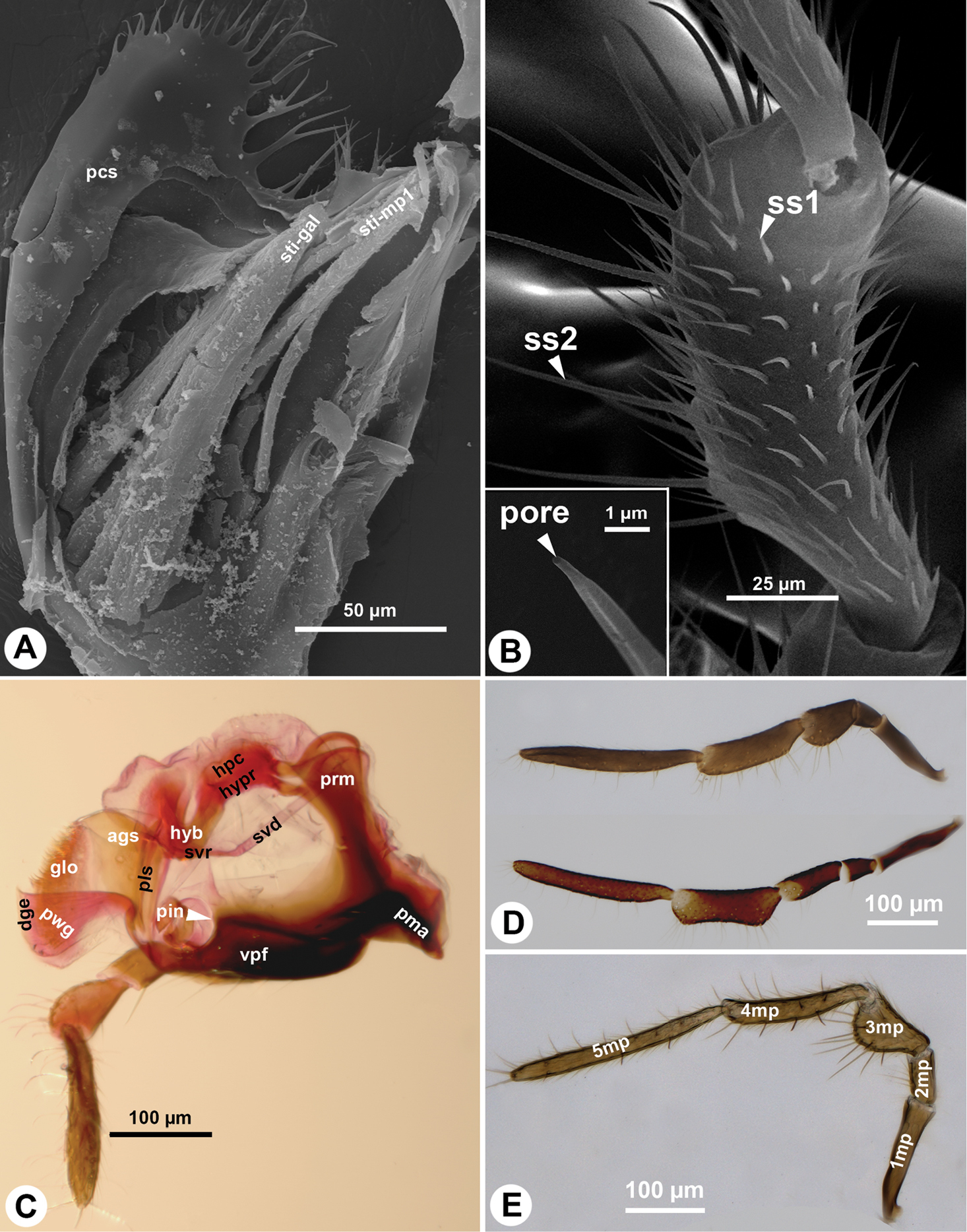
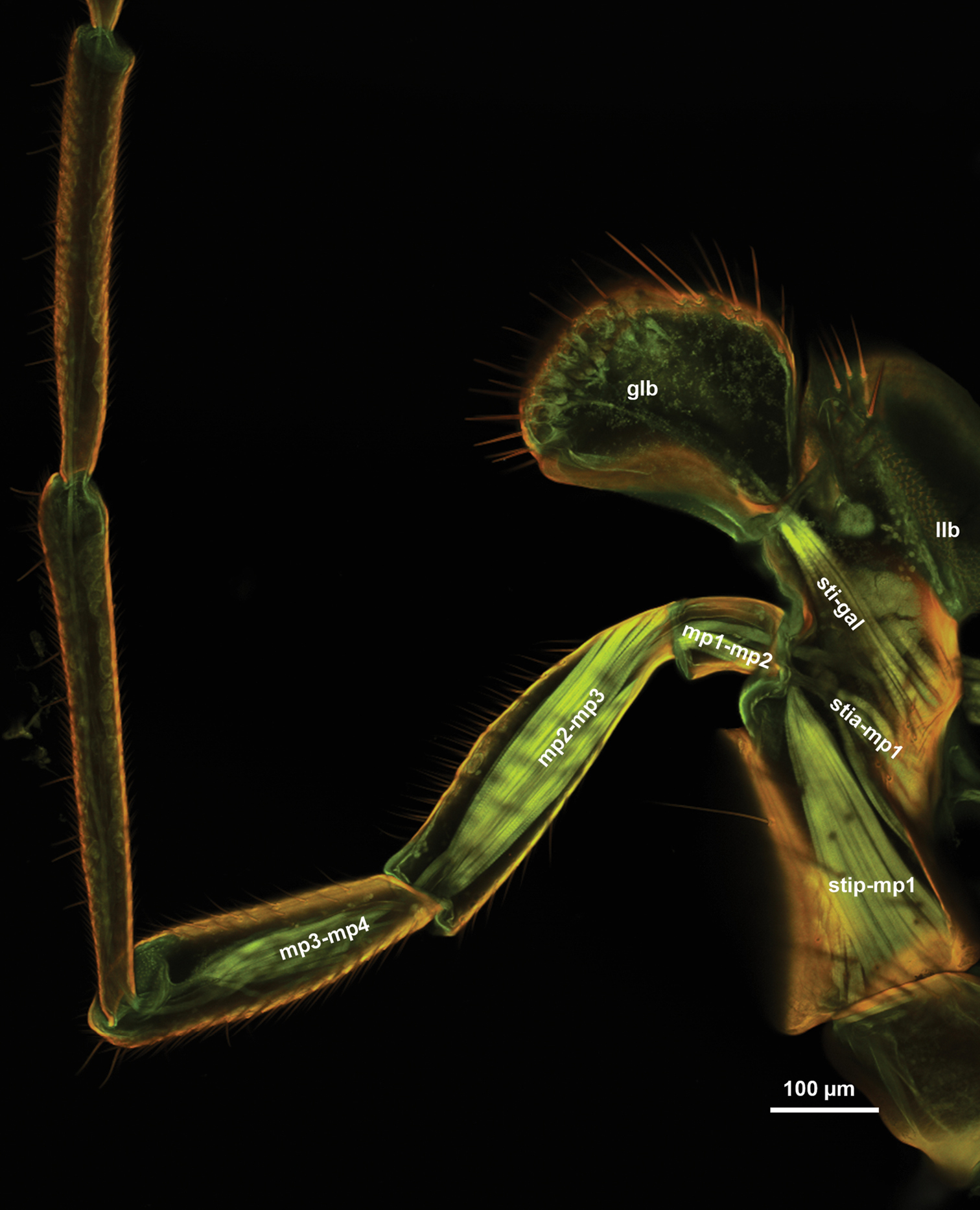
The maxillary palp (1mp: Fig. 2B, D) is connected at the distal apex of the posterior stipital sclerite trough the ventral dististipital process (vdp: Fig. 2B) adjacent to the basal galeal sclerite. The maxillary palp is composed of five maxillary sclerites among which the second sclerite of the maxillary palp is the shortest, and the fifth sclerite of the maxillary palp is always the longest (5mp: Figs 1A, C, 3D, E). The relative width of the maxillary palpal sclerites varies between Sparasion species and in some cases even between different sexes (Table 1). Two different setal types can be differentiated on the maxillary palp, based on their gross morphology. The type 1 seta (ss1: Fig. 3B) is a uniporous sensillum whereas the type 2 seta (ss2: Fig. 3B) is a longer mechanosensory hair. A type 1 seta is present on all but the first sclerite of the maxillary palp, which is glabrous in Sparasion sp. 4 and sp. 9 and bears only 1–2 type 2 setae in the rest of the species. Type 1 setae are evenly distributed on the third sclerite of the maxillary palp, on the fourth sclerite of the maxillary palp and on the fifth sclerite of the maxillary palp. Type 1 setae are located in 1–4 whorls of setae on the second sclerite of the maxillary palp. Type 2 setae occur only on the third, fourth sclerite of the maxillary palp and on the fifth sclerite of the maxillary palp. The number of type 2 setae is positively correlated with the width of the third sclerite of the maxillary palp and the fourth sclerite of the maxillary palp: type 2 setae are absent from the third sclerite of the maxillary palp if it is not increased in width relative to the second maxillary palpal sclerite.
The cranio-cardinal muscle (cr-cd: Figs 1B, 4A, B) arises medially of the posterior site of origins of the tentorium and inserts on the cardo just laterally of the cranial fossa of the cardo (caf: Fig. 1B). The tentorio-cardinal muscle (tnt-cd: Fig. 1B) arises ventrally on the tentorium just laterally of the site of origin of the tentorio-stipital muscle (tnt-sti: Fig. 4A) and inserts laterally on the stipitocardinal hinge. The tentorio-stipital muscle arises ventrally from the tentorium, medially of the site of origin of the tentorio-cardinal muscle and inserts on the median margin of the posterior stipital sclerite just posterior of the lacinial lever. The cranio-lacinial muscle is absent. The stipito-lacinial muscle (sti-lac: Fig. 2A) arises from along the lateral margin of the posterior stipital sclerite and inserts apically on the lacinial lever. The single stipito-palpal muscle (sti-mp1: Figs 2A, 3A) and the stipito-galeal muscle (sti-gal: Fig. 3A) arise medially from the stipito-lacinial muscle, subsequently. The stipito-galeal muscle inserts on the basal galeal sclerite. The first intrinsic muscle of the maxillary palp, third intrinsic muscle of the maxillary palp and fourth intrinsic muscle of the maxillary palp are present, the second intrinsic muscle of the maxillary palp was not observed. No muscle was attached to the fifth maxillary sclerite.
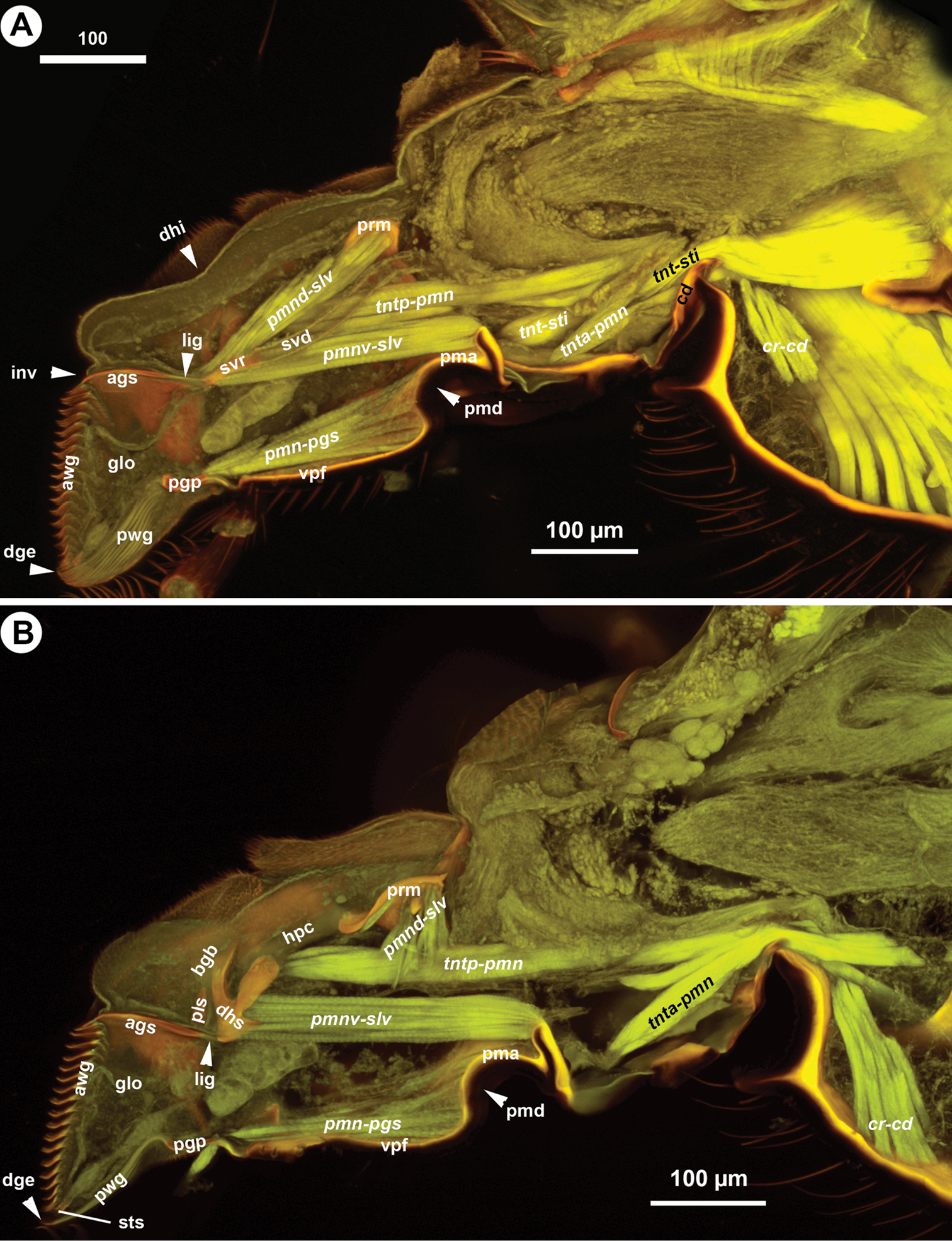
CLSM volume rendered images showing the skeletomuscular system of the labium of Sparasion, medial view, distal to the left.
The postmentum is composed of a single interstipital sclerite that is elongate, rectangular, and is articulated with the posterior stipital sclerites proximolaterally at the proximomedial stipital flange (psm, pmf: Figs 1B, C, 2C). The median postmental ridge (mpr: Figs 1A, B, 2C) is present and the postmental-premental hinge (pph: Figs 1C, 2C) is resilin rich.
The prementum is connected to the stipes via a conjunctiva (stipito-premental conjunctiva) extending along the proximal margin of the premental arms (prm: Figs 1B, 3C, 4A, B, 5C, 6A–D) and the lateral margin of the prementum proximally of the arm. The premental carinae (pmc: Fig. 2C and corresponding premental ditches (pmd: Figs 2C, 4A, B, 5C, D, 6A, D) converge proximally and separate the ventral premental face from (vpf: Figs 2C, 3C, 4A, B, 5C, D, 6A, D) the lateral premental face (pma: Figs 2C, 4A, B, 5C, D, 6A, D). The ventral premental face is flat, diamond-shaped and is equipped with campaniform sensillae of the prementum (cps: Fig. 2C) and distally-oriented mechanosensory hairs (msh: Fig. 2C). The number and pattern of both the campaniform sensilla and mechanosensory hairs and the length of the mechanosensory hairs are variable between different species and sexes (Table 1). The lateral face of the prementum is mostly overlapped ventrally by the distomedial stipital flange (Fig. 1A). The labial palpal excision (pin: Figs 2C, 3C) accommodating the base of the labial palp is distinct on the distolateral corner of the prementum. The labial palp is composed of three sclerites that are equipped with both the uniporous sensilla (type 1) and mechanosensory hairs (type 2).
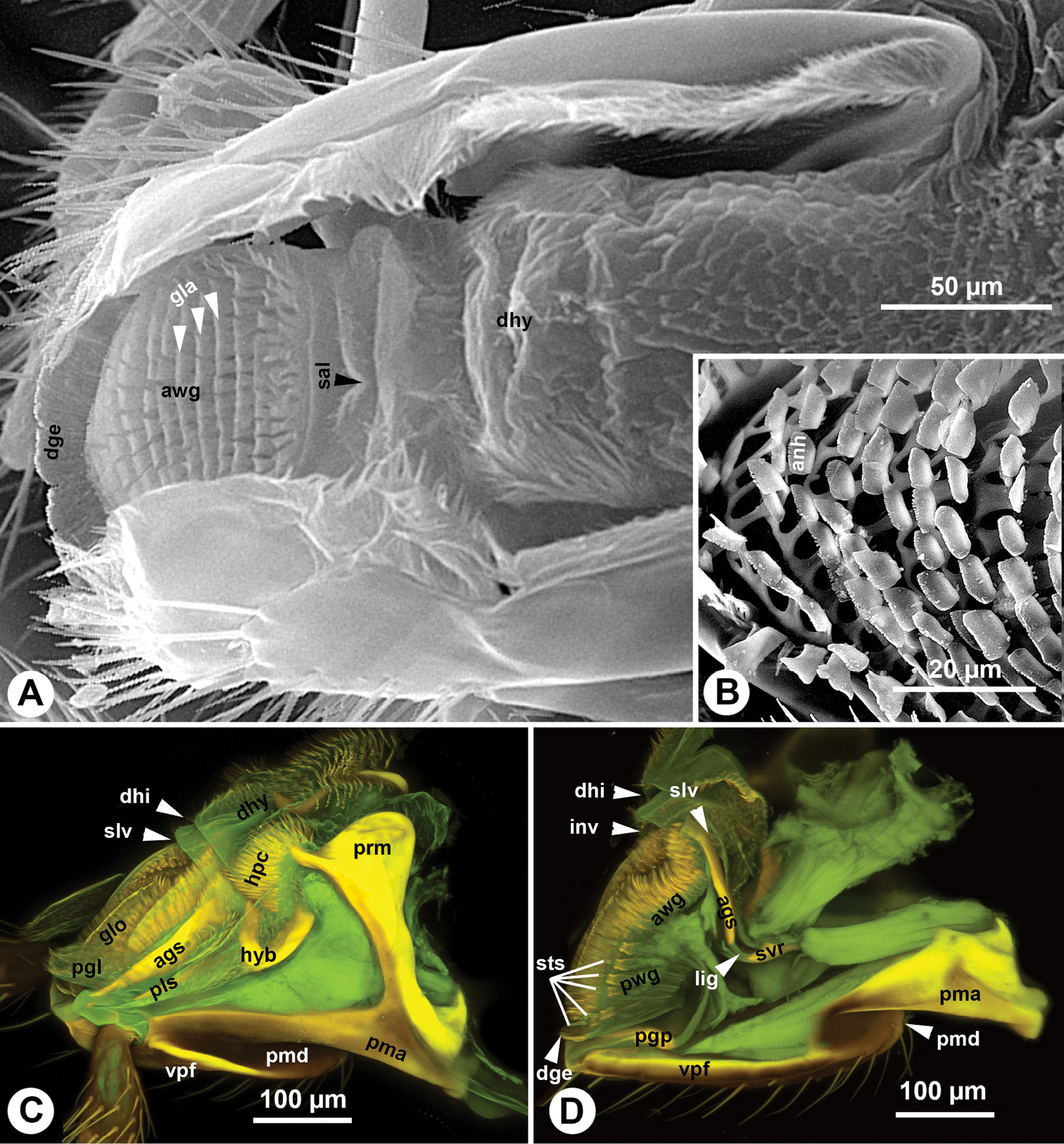
Labium and distal hypopharynx of Sparasion. A SEM micrograph showing the labium and distal hypopharynx, anterolateral view, distal to the left B SEM micrograph showing the glossal annuli, anterior view, distal to the left C, D CLSM volume rendered images showing the labium with retracted glossa and paraglossae, distal to the left C lateral view (doi: 10.6084/m9.figshare.861064) D medial view (doi: 10.6084/m9.figshare.861061).
Labium of Sparasion. A CLSM volume rendered images showing the labium with protracted glossa and paraglossae, lateral view, distal to the left (doi: 10.6084/m9.figshare.861063) B CLSM volume rendered images showing the labium, glossa removed, lateral view, distal to the left (doi: 10.6084/m9.figshare.861062, doi: 10.6084/m9.figshare.861066) C CLSM volume rendered images showing the labium, glossa removed, posterior view, distal to the left (doi: 10.6084/m9.figshare.861056, doi: 10.6084/m9.figshare.861067) D CLSM volume rendered images showing the labium with protracted glossa and paraglossae, medial view, distal to the left (doi: 10.6084/m9.figshare.861059) E, F Bright field images showing the labium, posterior view, distal to the top.
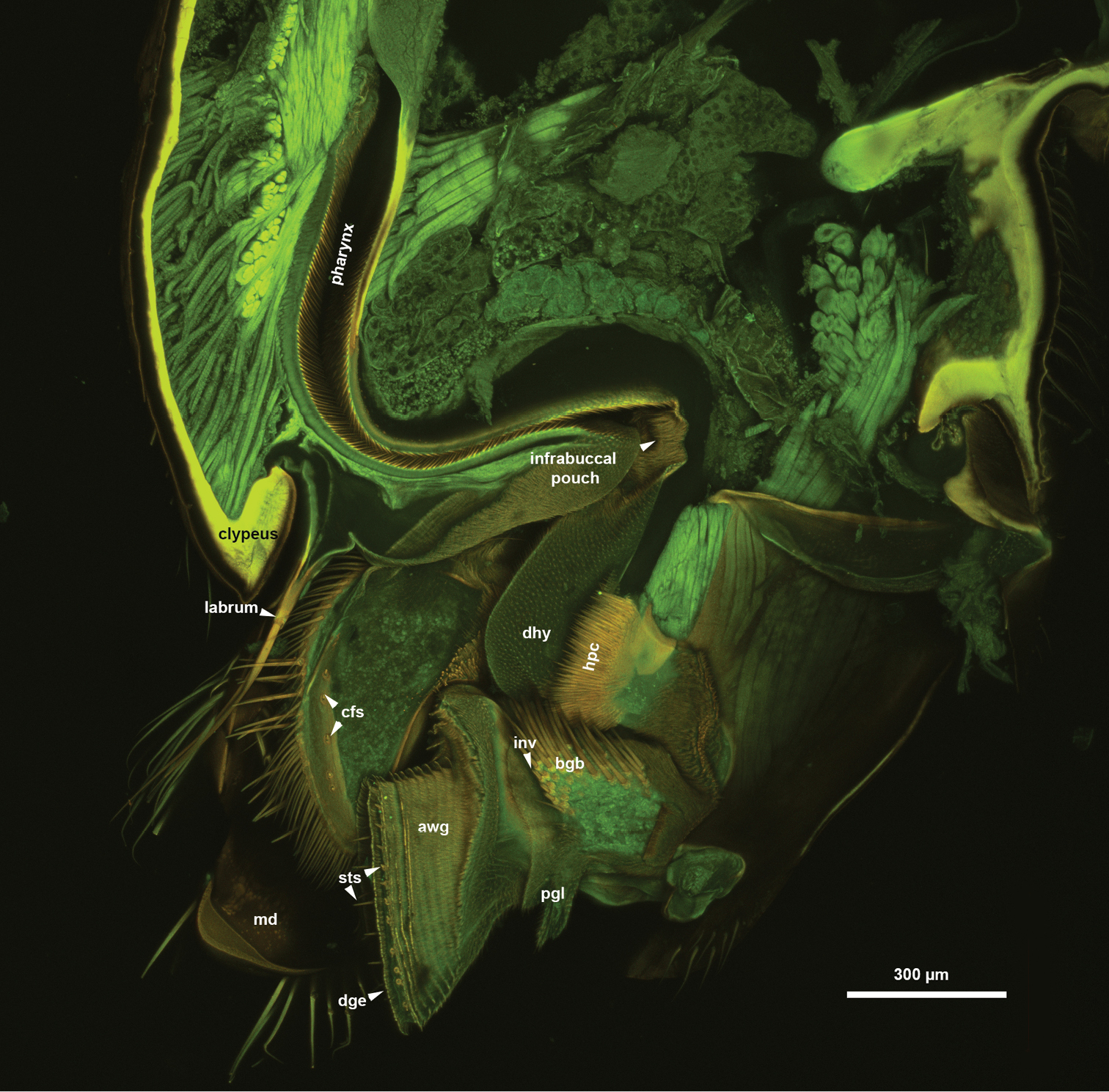
The glossa (glo: Figs 1C, 3C, 4A, B, 5C, 6A) is separated anteroproximally from the paraglossae (pgl: Figs 5C, 6A, C) and the distal hypopharynx by the basal glossal invagination (inv: Figs 4A, 5D, 6D) adjacent to the salivarium (svr: 6B, C, F). The anterior glossal sclerites (ags: Figs 3C, 4A, B, 5C, 6A, D) are embedded into the distal wall of the invagination and are connected medially to the ventral wall of the salivarium via a ligament (lig: Figs 4A, B, 5D, 6D). The ventrolateral basiglossal arms are distinct (vba: Fig. 6A, F) and each anterior glossal sclerite is equipped with a single campaniform sensillum of the glossa (csg: Fig. 6F). The posterior glossal plate is composed of a single, median sclerite (pgp: Figs 4A, B, 5D, 6E, F). The flat posterior surface of the glossa (pwg: Figs 3C, 4A, B, 5D, 6D) is glabrous with distinct ventral glossal lines. The distal glossal edge (dge: Figs 3C, 4A, B, 5A, D, 6D) is thicker than the proximal region of the posterior surface of the glossa. The apical glossal hairs are usually present (agh: Fig. 6A). The apical glossal setae (sts: Figs 5D, 6D, E) are styloconic sensilla with variable number in different species (Table 1).
The arched anterior surface of the glossa (awg: Figs 4A, B, 5A, D, 6D) is evenly covered with glossal annuli (gla: Fig. 5A), which are composed of transverse rows of spatulate and anteriorly curving annular hairs, each with a dentate distal margin (anh: Fig. 5B).
The bilobed paraglossae arise proximolaterally of the basal glossal invagination and encircle the proximal part of the glossa. The elongate, triangular basiparaglossal sclerite (pls: Figs 3C, 5C, 6A–D, F) corresponds to a posterior lobe of the paraglossa whereas the larger anterior lobe of the paraglossa bears the distal, less sclerotised paraglossal acroglossal button (pgb: Fig. 6B). Proximally the paraglossa is continuous with the wall of the distal hypopharynx (dhy: Fig. 5A, C) and the paraglossal sclerite is connected to the distal hypopharyngeal sclerite via an elongate ligament. The basiparaglossal brush (bgb: Figs 4B, 6A, B, F) is distinct but the paraglossal sclerite and paraglossal annuli are absent.
The distal hypopharynx is supported laterally by the premental arms and the hypopharyngeal rod (hypr: Fig. 3C), which is continuous with the distal hypopharyngeal sclerite via the hypopharyngeal button (hyb: Figs 3C, 5C, 6A–D). The hypopharyngeal pecten (hpc: Figs 4B, 5C, 6A, B) is distinct and abuts the basiparaglossal brush when the ligula is retracted (hpc: Figs 5C). The hypopharyngeal buttons are connected to each other medially trough the ventral wall of the salivarium, that is either sclerotised (svr: Figs 3C, 6B; salivarial sclerite) or is a resilin rich conjunctiva (svr: Fig. 6F). In the former case the hypopharyngeal buttons and the ventral, sclerotised wall of the salivarium compose the distal hypopharyngeal sclerite (dhs: Figs 4B, 6C) encompassing ventrally the salivary duct (svd: Figs 3C, 4A). The distal hypopharyngeal invagination (dhi: Figs 4A, 5C, D) corresponds to the dorsal bend on the hypopharyngeal button.
The postmento-premental muscle is absent. The posterior tentorio-premental muscle (tntp-pmn: Fig. 4A, B) arises from the tentorium anterolaterally from the site of origin of the anterior tentorio-premental muscle (tnta-pmn: Fig. 4A, B) and inserts on the proximal part of the hypopharyngeal button. The anterior tentorio-premental muscle arises from the tentorium medially of the site of origin of the posterior tentorio-premental muscle and inserts proximomedially on the postmental-premental hinge. The premento-paraglossal muscle (pmn-pgs: Fig. 4A, B) arises medially from the internal edge corresponding to the premental ditch and from the intima of the median premental face anterior to the edge and inserts on the posterior glossal plate. The dorsal premento-salivarial muscle (pmnd-slv: Fig. 4A, B) arises distomedially from the premental arm and inserts dorsally on the salivarium. The ventral premento-salivary sclerite muscle (pmnv-slv: Fig. 4A, B) arises from the proximal premental area and inserts proximomedially on the distal hypopharyngeal sclerite. The premento-palpal muscle arises anterolaterally from the site of origin of the premento-paraglossal muscle. The first intrinsic muscle of the labial palp and the second intrinsic muscle of the labial palp are present. The premento-glossal muscle is apparently absent.
The term maxillo-labial complex (MLC) is used by
The hymenopteran labium, unlike that of other holometabolan insects, is so tightly connected to the maxillae by the labiomaxillary hinge that they protract and retract together as a single unit (
Although the MLC of Sparasion is reduced in size and highly sclerotized, we were able to homologize most of its anatomical structures to those of other Hymenoptera (Appendix 2, Figs 7–9). We hypothesize that the platygastroid MLC is an anatomical system with enough phenotypic diversity to serve as a source of morphological characters for phylogenetic analyses and species diagnosis.
CLSM volume rendered images showing the MLC of Athalia rosae. A maxilla, medial view, doi: 10.6084/m9.figshare.956279 B MLC, anterior view, distal to the left, doi: 10.6084/m9.figshare.956279
CLSM volume rendered image showing the mouthparts of Evania sp., posterior view, distal to the bottom, doi: 10.6084/m9.figshare.956280
CLSM volume rendered image showing the maxilla of Orthogonalys pulchella, posteromedial view, distal to the top-left, doi: 10.6084/m9.figshare.956281
One consequence of the more heavily sclerotised and less moveable MLC is that the cardo remains relatively simple in Sparasion, retaining its main function: providing rigid attachment of the MLC to the cranium. The inner and outer processes of the cardo (
Different levels of sclerotization can be observed on the intercardinal and interstipital areas in different hymenopteran taxa. Only the intercardinal area contains a sclerite in basal Hymenoptera (
Only the interstipital postmentum (
The development of the principal carina arguably correlates with the presence of rigid connection between the stipes and neighboring sclerites. In those taxa, where the stipes articulates with the hypostoma, but not with the labium, the principal carina is present only laterally (Evania, pcs: Fig. 8).
Both the medial and lateral portions of the carina are well developed in Sparasion, having two unique characteristics that were not found in other Hymenoptera (see references listed in the materials and methods section):
1. The median portion of the carina is divided into two sections that overlap each other and accommodate the proximal and distal part of the lateral premental margin.
2. The presence of an “apical fringe”, a branched flattened evagination along the distolateral margin of the carina. While the first specialization might be the consequence of the articulation between the posterior stipital sclerite and the prementum, the second is most probably related to the unique movement of the mandible in Sparasion. While pivotal axis of the mandible is anteroposterior in most Hymenoptera taxa (dorsoventral in a prognathous head) it is directed mediolaterally in Sparasion and the mandibles are moved parallel to the body axis (
The presence of the campaniform sensillum of the stipes was considered as one of the synapomorphies of the clade composed of Platygastroidea and Cynipoidea (
Appendage segments and annuli are ring-like and repetitive sclerites of the legs, antenna, labial palps and maxillary palps. While appendage segments have muscles attaching to them, annuli do not. Annuli are traditionally differentiated from appendage segments by names with the suffix “-mere” e.g. flagellomere and tarsomere. Among the five maxillary palpal sclerites of Sparasion the fifth one does not have muscle attachments. We observed a similar condition in Orthogonalys (Trigonalidae), where the fifth and sixth sclerites lack any muscle attachment (Fig. 9). If we apply the annuli vs. segment terminology system to these taxa, we should call the first four sclerites “palpal segments” and fifth sclerite in Sparasion and the fifth and sixth sclerites in Orthogonalys “palpomeres”. To avoid misinterpretations of the segment identity of the palpi, and keeping a simple and easily applicable terminology we prefer to use palpal sclerites for the ring-like sclerites of the palpi.
Sparasion has five maxillary palpal sclerites that is the highest in Platygastroidea (
The maxillary palp is located medially on the lateral margin of the posterior stipital sclerite in basal Hymenoptera distantly from the apical end of the sclerite bearing the base of the galea (Fig. 7B,
The 2nd, 3rd and 4th maxillary palpal sclerites are distinctly wider than the more proximal or distal maxillary palpal sclerites in some Sparasion species (Figures 3D, E). The number and position of these modified sclerites are sexually dimorphic in some Sparasion species and are seemingly useful for species groups definitions. A widened 4th palpal sclerite was found in most species of the plesiomorphic platygastrid genus Nixonia, and representatives of Plaumannion, Archaeoteleia, Sceliomorpha and Neuroscelio (
Although the position of enlarged palpal sclerites in the maxillary palp has often been used for the classification of different Hymenoptera taxa (i.e. Evaniidae,
Although the presence of only two stipito-palpal muscles is considered as the Hymenoptera groundplan (stia-mp1, stip-mp1: Fig. 9;
The terms, galea and lacinia refer usually to the evaginated distal, sclerotised regions of the maxillar cuticle that are adjacent to the sites of insertions for the stipito-lacinial, cranio-lacinal and the stipito-galeal muscles in insects. Although these muscles are present in most hymenopterans (sti-gal, sti-lac: Fig. 7A, and more proximal and more distal lobes can almost always be differentiated, the proximal limits of the galea and lacinia are difficult to define and thus these structures are difficult to homologize to that of other insects where the galea and lacinia are well sclerotised. Therefore we preferred to use galeal lobe and lacinial lobe instead of galea and lacinia in the present paper.
Two distinct setiferous areas can be defined on the lateral area of the galeal lobe in Athalia: a proximal row of setae traversing the galea (prs: Fig. 7B) and a more distal setiferous patch (dsp: Fig. 7B). The presence of these two setiferous areas seems to be consistent within Hymenoptera (
In Sparasion, similarly to some other, more derived Hymenoptera taxa these setiferous areas correspond to two sclerites, the proximolateral and distolateral sclerites of the galea (
Although the velum is well developed in numerous other Hymenoptera (
The galeal comb and the galeal lamina are important characters that have been used in aculeate systematics (
The prementum is articulated with the stipes via the premento-stipital articulation that is composed of the premental ditch, premental carinae, and the principal carina of the stipes.
CLSM volume rendered image showing the ventral region of the head of Evania sp., medial view, distal to the bottom, doi: 10.6084/m9.figshare.956282
The maximum number of labial palpal sclerites in Apocrita is four (some Formicidae (Gotwald, 1969), Braconidae (Belokobylskij, 2006), Vespinae (Duncan, 1939)). In Sparasion, the labial palp is composed of three sclerites that are moved directly by muscles. In those Hymenoptera where the labial palp is composed of four sclerites (Vilhelmsen and Beutel 2007,
Along the proximal margin of distal edge of glossa in Sparasion, a row of 9–23 apical glossal setae can be observed. These setae are styloconic sensilla, which are considered to be bimodal contact chemo-mechanosensillae (
The ventral glossal lines have been reported only from a few Hymenoptera (
We wish to thank Mariana Popovici and Gordon Ramel for gift specimens, Irina Gostin and Răileanu for the SEM pictures, Eva Johannes (NCSU-NSCORT, Department of Plant Biology) for her assistance in Confocal Laser Scanning Microscopy, Lubomír Masner, Lucian Fusu, Mircea Mitroiu, and Lars Vilhelmsen for their valuable comments, and Norman F. Johnson and Luciana Musetti for maintaining the sites “The genera of Platygastroidea (Hymenoptera) of the World” and “Hymenoptera On-line Database”. The material is based on work supported, in part, by a grant of the Romanian National Authority for Scientific Research, CNCS-UEFISCDI, project number PN-II-RU-TE-2012-0057 to O. Popovici. This research is also based on work supported by the National Science Foundation (grant numbers DBI-0850223, DEB-0842289, DEB-0956049, and EF-0905606); and the National Evolutionary Synthesis Center and benefited from discussions initiated through the Phenotype Research Coordination Network. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation.
Specimens examined.
| Taxon | Number of specimens | Data labels |
|---|---|---|
| Anteris sp. | 1 | ROMANIA: Botosani, 10.viii.2004, leg. O. Popovici |
| Anteris sp. | 1 | ROMANIA: Iasi, 26.vi.2004, leg. O. Popovici |
| Apegus sp. | 1 | ROMANIA: Iasi, 30.vi.2004, leg. O. Popovici |
| Athalia rosae | 1 | Wildtype culture, 2005, M. Hatakeyama |
| Belyta sp. | 2 | ROMANIA: Iasi, 10.ix.2005, leg. O. Popovici |
| Calliscelio sp. | 1 | ROMANIA: Bacău, 31.viii.2002 , leg. O. Popovici |
| Calliscelio sp. | 1 | ROMANIA: Holboca (Iaşi), 30.06.2002, leg. I. Moglan |
| Duta sp. | 1 | ROMANIA: Bacau, 17.vii.2004, leg. O. Popovici |
| Evania sp. | 2 | Costa Rica, 21.II.2005 15/M/16/016 1500-site Costa Rica Heredia, 9km NE Vara Blanca, 10°14'N (INBIO) |
| Gasteruption sp. | 2 | USA WV, Hardy Co., 11–22.v.2006, MT, D. Smith |
| Ibalia sp. | 1 | USA: Virginia, Fairfax Co., Great Falls, 18–30.v. 2007, leg. D. Smith |
| Ibalia sp. | 1 | USA: Motgomery Co., 21.v. 2010, leg. N. E. Woodley |
| Ibalia sp. | 1 | USA: Virginia, Fairfax Co., Great Falls, 31.v.-13.vi.2007, leg. D. Smith |
| Macroteleia sp. | 1 | ROMANIA: Iasi, 17.viii.2004, leg. O. Popovici |
| Macroteleia sp. | 1 | ROMANIA: Iasi, 13.viii.2000, leg. M. Mitroiu |
| Macroteleia sp. | 1 | ROMANIA: Iasi, 25.vii.2005, leg. L. Fusu |
| Orthogonalys pulchella | 4 | USA, WV, Hardy Co. 3mi NE Mathias, 38°55'N, 78°49'W, 2–15.vii.2004, Malaise trap. D. Smith |
| Paramesius sp. | 1 | ROMANIA: Iasi, 10.ix.2005, leg. O. Popovici |
| Pelecinus polyturator | 1 | USA: VA: Fairfax Co., Great Falls, 27.vii–4.viii. 2006, leg. D. Smith |
| Pelecinus polyturator | 1 | USA: VA: Fairfax Co., Turkey Run, 5–24.viii.2006, leg. D. Smith |
| Proctotrupes sp. | 1 | GREECE: Kerkini Nat. Res., 15–17.vii.2007, leg. G. Ramel |
| Proctotrupes sp. | 1 | ROMANIA: Iasi, 7.x.2004, leg. O. Popovici |
| Proctotrupes sp. | 1 | ROMANIA: Vorona, 21.v.2007, leg. C. Lisenchi |
| Psilanteris sp. | 1 | ROMANIA: Bacau, 17.vii.2004, leg. O. Popovici |
| Ropronia sp. | 2 | USA: Virginia, Fairfax Co., 18–30.v.2007, leg. D. Smith |
| Scelio sp. | 3 | ROMANIA: Iasi, 24.viii.2004, leg. O. Popovici |
| Sparasion sp.1 | 2 | ROMANIA: Iasi county, Bârnova forest, meadow in glade; 12.vii.2007 N46°59.617', E27°35.452', Leg. Popovici O. & Popovici Mariana YPT |
| Sparasion sp.2 | 1 | ROMANIA: Iasi county, Bârnova forest, meadow in glade; 12.vii.2007 N46°59.617', E27°35.452', Leg. Popovici O. & Popovici Mariana YPT |
| Sparasion sp.2 | 2 | ROMANIA: Iasi county, Botanical Garden; N47°11.167', E27°32.983'; 26.vii.2007; Leg. Popovici O. YPT |
| Sparasion sp.3 | 3 | ROMANIA: Constanta county; sand dune natural reservation from Agigea; 27–29.vii.2008; N44°5.184', E28°38.517'; Leg. Popovici O. & Popovici Mariana YPT |
| Sparasion sp.4 | 10 | ROMANIA: Constanta county; sand dune natural reservation from Agigea; 27–29.vii.2008; N44°5.184', E28°38.517'; Leg. Popovici O. & Popovici Mariana YPT |
| Sparasion sp.5 | 1 | ROMANIA: Tulcea county; forest border, 9 km S of Babadag; 6–7.vii.2009; N44°48.817', E28°42.41'; Leg. Popovici O. & Fusu L. YPT |
| Sparasion sp.6 | 1 | ROMANIA: Tulcea county; forest border, 9 km S of Babadag; 6–7.vii.2009; N44°48.817', E28°42.41'; Leg. Popovici O. & Fusu L. YPT |
| Sparasion sp.7 | 1 | ROMANIA: Tulcea county; forest border, 9 km S of Babadag; 6–7.vii.2009; N44°48.817', E28°42.41'; Leg. Popovici O. & Fusu L. YPT |
| Sparasion sp.7 | 2 | GREECE: Kerkini Lake, N.P. Lithotopos, 1–7.viii.2006 leg. G. Ramel |
| Sparasion sp.8 | 2 | JAPAN: Kitahira, Otsu-shi Shiga-ken, 19–22.vii.2008, leg. T. Yoshida. |
| Sparasion sp.9 | 1 | UGANDA: Kibale, N.P. Kanyawara Biol. Station, 14–21.xi.2010, leg. S. Katusabe & Co. |
| Sparasion sp. | 1 | USA: NC, Raleigh 03.VII.2009 Winkler, Benoit |
| Sparasion sp. | 7 | USA: West Virginia, Hardy Co. VIII-11–28–06, D. Smith |
| Triteleia sp. | 1 | ROMANIA: Iasi, 8.ix.2004, leg. O. Popovici |
| Vanhornia sp. | 3 | USA: VA: Loudoun Co., 1.vi–17.vii.2000, leg. D. Smith |
Anatomical terms used, cross-referenced to an ontological (formal) definition (Hymenoptera Anatomy Ontology; URI = Uniform Resource Identifier).
| Abbreviation | Term | Ontological definiton | URI |
|---|---|---|---|
| acanthae | The process that corresponds to a single epidermal cell. | http://purl.obolibrary.org/obo/HAO_0002119 | |
| anh | annular hair | The spine on the distal part of ligula that is connected proximolaterally to the neighboring glossal hair by a carina. | http://purl.obolibrary.org/obo/HAO_0002205 |
| ags | anterior glossal sclerite | The sclerite of the anterior glossal plate that is lateral to the median conjunctiva of the anterior glossal plate. | http://purl.obolibrary.org/obo/HAO_0000112 |
| asg | anterior surface of the glossa | The area of the glossa that is between the distal glossal edge and the anterior glossal plate. | http://purl.obolibrary.org/obo/HAO_0002208 |
| tnta-pmn | anterior tentorio-premental muscle | The muscle that arises anteriorly from the cranium and inserts on the posterior margin of the prementum. | http://purl.obolibrary.org/obo/HAO_0001064 |
| stia-mp1 | anterior stipito-palpal muscle | The stipito-palpal muscle that arises medially on the proximal part of the stipes and inserts anteroproximally on the first maxillary sclerite. | http://purl.obolibrary.org/obo/HAO_0000909 |
| conj | anterolateral wall of the stipes | The area that extends between anterior margin of the median wall of the stipes and the lateral margin of the posterior wall of the stipes and the base of the mandible and the labrum. | http://purl.obolibrary.org/obo/HAO_0002215 |
| agh | apical glossal hair | The spines that are on the distal glossal edge. | http://purl.obolibrary.org/obo/HAO_0002209 |
| sts | apical glossal setae | The row of setae of the glossa that is adjacent to the distal glossal edge. | http://purl.obolibrary.org/obo/HAO_0002210 |
| apicomedial galeal plate | The sclerite that is located on the medial surface of the galeo-lacinial complex and bears the coeloconic sensillum of galea. | http://purl.obolibrary.org/obo/HAO_0002142 | |
| bcr | basal cardinal ridge | The cardinal ridge that is median. | http://purl.obolibrary.org/obo/HAO_0002083 |
| bgs | basal galeal sclerite | The sclerite that receives the site of insertion of the stipito-galeal muscle. | http://purl.obolibrary.org/obo/HAO_0002143 |
| inv | basal glossal invagination | The anterior invagination of the labium that is adjacent with the distal end of the salivarium and separates the glossa proximally from the distal hypopharynx and the paraglossae. | http://purl.obolibrary.org/obo/HAO_0002231 |
| bgb | basiparaglossal brush | The anteroproximal area of the paraglossa that is covered with long acanthae and corresponds to the ligament connecting the acroglossal button with the basiparaglossal sclerite. | http://purl.obolibrary.org/obo/HAO_0002199 |
| pls | basiparaglossal sclerite | The sclerite that is located medially on the paraglossa and articulates with the anterior glossal plate and is continuous with the hypopharyngeal rod.. | http://purl.obolibrary.org/obo/HAO_0002201 |
| cps | campaniform sensillum | The aporous sensillum without a hairlike cuticular component. | http://purl.obolibrary.org/obo/HAO_0001973 |
| cfs | coeloconic sensillum of galea | The coeloconic sensillum that is located on the medial surface of the galeo-lacinial complex distal to the base of the lacinial lobe. | http://purl.obolibrary.org/obo/HAO_0002141 |
| csg | campaniform sensillum of glossa | The campaniform sensillum of the anterior surface of the glossa that is located proximal to the glossal annuli. | http://purl.obolibrary.org/obo/HAO_0002212 |
| cps | campaniform sensillum of the prementum | The campaniform sensillum that is on the ventral premental face. | http://purl.obolibrary.org/obo/HAO_0002244 |
| cardinal articular condyle of the cranium | The condyle that is located on the cranium and articulates with the cranial fossa of the cardo. | http://purl.obolibrary.org/obo/HAO_0002074 | |
| cardinal lever | The process that receives the site of attachment of the cranio-cardinal muscle. | http://purl.obolibrary.org/obo/HAO_0002075 | |
| cd | cardo | The sclerite that is articulated with the cranium at the cranio-cardinal articulation, is connected to the stipes distolaterally via the stipitocardinal hinge and receives the site of attachment of the cranio-cardinal muscle. | http://purl.obolibrary.org/obo/HAO_0000187 |
| conjunctiva | The area of the integument that is weakly sclerotized, with thin exocuticle. | http://purl.obolibrary.org/obo/HAO_0000221 | |
| caf | cranial fossa of the cardo | The fossa that is located on the cardo and articulates with the cardinal condyle of the cranium. | http://purl.obolibrary.org/obo/HAO_0002219 |
| cr-cd | cranio-cardinal muscle | The maxillar muscle that arises medially from the occiput dorsally of the occipital foramen and inserts on the proximolateral part of the cardo. | http://purl.obolibrary.org/obo/HAO_0001592 |
| cranio-lacinial muscle | The maxillar muscle that arises from the occiput and inserts on the proximal part of the lacinial lobe. | http://purl.obolibrary.org/obo/HAO_0001593 | |
| dge | distal glossal edge | The transverse edge that extends distally on the glossa. | http://purl.obolibrary.org/obo/HAO_0002206 |
| dhi | distal hypopharyngeal invagination | The invagination on the distal hypopharynx that id adjacent to the concavities of the hypopharyngeal rods. | http://purl.obolibrary.org/obo/HAO_0002213 |
| dhy | distal hypopharynx | The area that is located on the anterior surface of the hypopharyngeal wall and is delimited proximally by the infrabuccal pouch or the distal margin of the sitophore, distally by the salivarial orifice and laterally by the lateral parts of the prementum and the hypopharyngeal rods. | http://purl.obolibrary.org/obo/HAO_0001575 |
| dhs | distal hypopharyngeal sclerite | The sclerite that receives the site of insertions of the dorsal and ventral premento-salivarial muscles and is continuous with the hypopharyngeal rod. | http://purl.obolibrary.org/obo/HAO_0002228 |
| dgs | distolateral galeal sclerite | The sclerite that is located distolaterally on the lateral wall of the galeo-lacinial complex and bears the apicolateral galeal setae. | http://purl.obolibrary.org/obo/HAO_0002127 |
| dgp | distolateral galeal setiferous patch | The setiferous patch that is located distolaterally on the galeo-lacinial complex | http://purl.obolibrary.org/obo/HAO_0002128 |
| dmf | distomedial stipital flange | The medial part of the principal carina of the stipes that is overlapped by the premental carina and overlaps the lateral premental face. | http://purl.obolibrary.org/obo/HAO_0002217 |
| pmd-slv | dorsal premento-salivarial muscle | The salivarial muscle that inserts proximodorsally on the salivary duct. | http://purl.obolibrary.org/obo/HAO_0000274 |
| 5mp | fifth sclerite of the maxillary palp | The sclerite that is ringlike and is connected distally to the fourth sclerite of the maxillary palp via conjunctiva. | http://purl.obolibrary.org/obo/HAO_0002220 |
| mp1-mp2 | first intrinsic muscle of the maxillary palp | The muscle that arises from the first sclerite of the maxillary palp and inserts on the second sclerite of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0002114 |
| first intrinsic muscle of the labial palp | The muscle that arises from the first sclerite of the labial palp and inserts on the second sclerite of the labial palp. | http://purl.obolibrary.org/obo/HAO_0002237 | |
| 1lp | first sclerite of the labial palp | The sclerite that is ringlike and is connected distolaterally to the prementum via conjunctiva and muscle. | http://purl.obolibrary.org/obo/HAO_0002194 |
| 1mp | first sclerite of the maxillary palp | The sclerite that is ringlike and is connected distolaterally to the posterior stipital sclerite via conjunctiva and muscle. | http://purl.obolibrary.org/obo/HAO_0002109 |
| fourth intrinsic muscle of the maxillary palp | The muscle that arises from the third sclerite of the maxillary palp and inserts on the fourth sclerite of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0002222 | |
| 4mp | fourth sclerite of the maxillary palp | The sclerite that is ringlike and is connected distally to the third sclerite of the maxillary palp via conjunctiva. | http://purl.obolibrary.org/obo/HAO_0002113 |
| galeal comb | The row of setae that is located on the medial wall of the galeo-lacinial complex proximal to the coeloconic sensilla of galea. | http://purl.obolibrary.org/obo/HAO_0002243 | |
| galeal fringe | The row of setae that extends along the margin of the galeo-lacinial complex, distal to the lacinial lobe. | http://purl.obolibrary.org/obo/HAO_0002133 | |
| galeal lamina | The lobe that is located medially on the internal wall of the galeo-lacinial complex and margined by the galeal comb. | http://purl.obolibrary.org/obo/HAO_0002136 | |
| glb | galeal lobe | The lobe that is located on the stipes at the distal part of the posterior stipital sclerite distolateral to the lacinia. | http://purl.obolibrary.org/obo/HAO_0000368 |
| gal-lac | galeo-lacinial complex | The area of the stipes that is delimited proximomedially by the stipito-premental conjunctiva, proximolaterally by the stipito-mandibular conjunctiva and posteroproximally by the margin of the posterior stipital sclerite. | http://purl.obolibrary.org/obo/HAO_0002126 |
| glo | glossa | The lobe of the labium that is limited posteroproximally by the prementum, anteroproximally by the fold traversing the salivary orifice and laterally by the paraglossae. | http://purl.obolibrary.org/obo/HAO_0000376 |
| gla | glossal annuli | The anatomical cluster of the glossa that is composed of annular hairs. | http://purl.obolibrary.org/obo/HAO_0002204 |
| hyb | hypopharyngeal button | The bent area of the hypopharyngeal rod that receives the site of insertion of the posterior tentorio-premental muscle. | http://purl.obolibrary.org/obo/HAO_0002234 |
| hpc | hypopharyngeal pecten | The anterolateral area of the distal hypopharynx that is adjacent with the proximal part of the hypopharyngeal rod proximal to the hypopharyngeal button and is covered with acanthae. | http://purl.obolibrary.org/obo/HAO_0002214 |
| hypr | hypopharyngeal rod | The ligament that connects the proximolateral margin of the prementum with the proximal part of the ligula. | http://purl.obolibrary.org/obo/HAO_0000408 |
| hys | hypostoma | The area that extends on the posterior (ventral) margin of the oral foramen along the site of attachments of the conjunctiva connecting the cranium with the maxillae and is delimited laterally by the pleurostomal fossa. | http://purl.obolibrary.org/obo/HAO_0000411 |
| integument | The anatomical system that forms the covering layer of the animal, ectodermal in origin and composed of epidermal cells producing the cuticle. | http://purl.obolibrary.org/obo/HAO_0000421 | |
| intercardinal area | The area that is located between the cardines and limited proximally and distally by the anatomical line that extends between the proximal and distal ends of the left and right cardines. | http://purl.obolibrary.org/obo/HAO_0002145 | |
| interstipital area | The area of the postmental area that is limited laterally by the median margins of the stipites, proximally by the anterior margin of the intercardinal area and distally by the proximal margin of the prementum. | http://purl.obolibrary.org/obo/HAO_0002146 | |
| interstipital sclerite | The sclerite that is located on the interstipital area and is connected to the prementum via conjunctiva. | http://purl.obolibrary.org/obo/HAO_0002223 | |
| invagination | The area where the cuticle is invaginated. | http://purl.obolibrary.org/obo/HAO_0002021 | |
| labial palp | The anatomical structure that is distal to the proximal margin of the first sclerite of the labial palp. | http://purl.obolibrary.org/obo/HAO_0000450 | |
| pin | labial palpal excision | The notch that is located distolaterally on the prementum and receives the base of the labial palp. | http://purl.obolibrary.org/obo/HAO_0002153 |
| labium | The appendage that is encircled by the area that is proximally delimited by the lateral margins of the cardo and the posterior stipital sclerite laterally, and the anatomical line that is tangential to the salivary duct and traverses the salivary orifice anteriorly. | http://purl.obolibrary.org/obo/HAO_0000453 | |
| lacinial bar | The sclerite that is located on the lateral wall of the lacinial lobe. | http://purl.obolibrary.org/obo/HAO_0002117 | |
| lacinial comb | The row of setae on the lacinial lobe that is marginal. | http://purl.obolibrary.org/obo/HAO_0002124 | |
| bls | lacinial lever | The sclerite that is located on the medial stipital wall and receives the site of insertion of the stipito-lacinial muscle. | http://purl.obolibrary.org/obo/HAO_0002093 |
| llb | lacinial lobe | The lobe that extends proximally on the distal margin of the medial stipital wall, is adjacent to the basal lacinial sclerite, overlaps the proximal part of the galea. | http://purl.obolibrary.org/obo/HAO_0000457 |
| pma | lateral premental face | The area of the prementum, that lays parallel and connected via conjunctiva to the medial stipital vall. | http://purl.obolibrary.org/obo/HAO_0002152 |
| lcr | laterodistal cardinal ridge | The cardinal ridge that arises distally from the basal cardinal ridge and is oriented distolaterally. | http://purl.obolibrary.org/obo/HAO_0002084 |
| ligament | The area of the cuticle that is resilin rich. | http://purl.obolibrary.org/obo/HAO_0002229 | |
| ligula | The anatomical cluster that is composed of the glossa and paraglossae. | http://purl.obolibrary.org/obo/HAO_0000496 | |
| lobes | The evagination that is mostly membranous. | http://purl.obolibrary.org/obo/HAO_0001587 | |
| md | mandible | The sclerite that is connected to the cranium along the anterior margin of the oral foramen via the anterior and posterior cranio-mandibular articulations. | http://purl.obolibrary.org/obo/HAO_0000506 |
| mfs | marginal fringe of the stipes | The anatomical cluster that is composed of the spines on the distal margin of the distolateral portion of the principal carina of the stipes. | http://purl.obolibrary.org/obo/HAO_0002216 |
| maxilla, maxillae | The appendage that is encircled by the area that is proximally delimited by the hypostoma posteriorly, the median margin of the mandible laterally, the labrum anterolaterally and the labium medially. | http://purl.obolibrary.org/obo/HAO_0000513 | |
| mpalp | maxillary palp | The anatomical structure that is distal to the proximal margin of the first sclerite of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0000515 |
| maxillary palpal sclerite | The sclerite that is part of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0002183 | |
| maxillary process of the hypostoma | The articular process that bears the cardinal condyle of the cranium. | http://purl.obolibrary.org/obo/HAO_0002073 | |
| maxillo-labial complex | The anatomical cluster that is composed of the maxillae and the labium and is connected by conjunctivae laterally to the cranium along the hypostoma, to the mandible along the proximomedial margin of the mandible and proximally to the hypopharynx. | http://purl.obolibrary.org/obo/HAO_0000452 | |
| msh | mechanosensory hair | The aporous sensillum that has a hair-like cuticular component. | http://purl.obolibrary.org/obo/HAO_0001039 |
| msg | medial stipital groove | The depression that extends medially and adjacent to the medial portion of the principal carina of stipes. | http://purl.obolibrary.org/obo/HAO_0002106 |
| medial wall | The area that is medial and lays parallel with the lateral wall of the prementum. | http://purl.obolibrary.org/obo/HAO_0002092 | |
| median conjunctiva of the anterior glossal plate | The median conjunctiva of the anterior glossal plate that is parallel with the anteroposterior body axis. | http://purl.obolibrary.org/obo/HAO_0002203 | |
| mpr | median postmental ridge | The ridge that is limited laterally by the stipital articular surfaces of the postmentum. | http://purl.obolibrary.org/obo/HAO_0002225 |
| mcr | mediodistal cardinal ridge | The cardinal ridge that arises distally from the basal cardinal ridge and is oriented distomedially. | http://purl.obolibrary.org/obo/HAO_0002085 |
| pgl | paraglossa | The lobe that is connected to the distal margin of the prementum posteroproximally, to the premental hypopharynx proximolaterally and anteroproximally, to the glossa proximomedially and bears the basiparaglossal brush and the paraglossal sclerite. | http://purl.obolibrary.org/obo/HAO_0000686 |
| pgb | paraglossal acroglossal button | The sclerite that is located distally on the paraglossa. | http://purl.obolibrary.org/obo/HAO_0002232 |
| paraglossal annuli | The anatomical cluster of the paraglossa that is composed of connected annular hairs. | http://purl.obolibrary.org/obo/HAO_0002233 | |
| paraglossal sclerite | The anteroproximal sclerite of the paraglossa that bears the basiparaglossal brush. | http://purl.obolibrary.org/obo/HAO_0002200 | |
| pgp | posterior glossal plate | The sclerite that is connected to the distal margin of the prementum and receives the site of insertion of the premento-paraglossal muscles. | http://purl.obolibrary.org/obo/HAO_0000747 |
| pss | posterior stipital sclerite | The sclerite that is located on the posterior stipital wall, articulates with the cardo and with the labial palp, is connected by conjunctiva distolaterally to the galeo-lacinial complex, proximally to the hypostoma and the cardo, proximolaterally to the mandible. | http://purl.obolibrary.org/obo/HAO_0002097 |
| stip-mp1 | posterior stipito-palpal muscle | The stipito-palpal muscle that arises medially on the proximal part of the stipes and inserts posteroproximally on the first maxillary palpal segment. | http://purl.obolibrary.org/obo/HAO_0001814 |
| posterior wall | The area that is limited medially by the medial stipital wall and limited proximolaterally by the margin of the posterior stipital sclerite. | http://purl.obolibrary.org/obo/HAO_0002097 | |
| pwg | posterior surface of the glossa | The area of the glossa that is between the distal margin of the prementum and the distal glossal edge. | http://purl.obolibrary.org/obo/HAO_0002207 |
| tntp-pmn | posterior tentorio-premental muscle | The tentorio-labial muscle that arises from the cranium and inserts distally on the labium adjacent to the level of the salivary orifice. | http://purl.obolibrary.org/obo/HAO_0000264 |
| postmental area | The area that is limited distally by the posterior margin of the prementum and laterally by the median margins of the cardines and the stipites. | http://purl.obolibrary.org/obo/HAO_0002144 | |
| pph | postmental-premental hinge | The conjunctiva that is between the postmentum and prementum. | http://purl.obolibrary.org/obo/HAO_0002226 |
| postmento-premental muscle | The labial muscle that is unpaired, arises from the postmentum and inserts on the proximal margin of the prementum. | http://purl.obolibrary.org/obo/HAO_0000803 | |
| psm | postmentum | The anatomical cluster that is composed of the sclerites that are on the postmental area. | http://purl.obolibrary.org/obo/HAO_0000785 |
| prm | premental arms | The area of the lateral premental face that receives the site of origin of the dorsal premento-salivarial muscle. | http://purl.obolibrary.org/obo/HAO_0002155 |
| pmc | premental carina | The flange that is adjacent with the border between the ventral and lateral premental faces and that overlaps externally the median part of the posterior stipital sclerite. | http://purl.obolibrary.org/obo/HAO_0002157 |
| pmd | premental ditch | The scrobe of the prementum that is adjacent to and extends lateral to the premental carina and accommodates the medial part of the posterior stipital sclerite. | http://purl.obolibrary.org/obo/HAO_0002227 |
| premento-glossal muscle | The labial muscle that arises on the ventral part of the prementum, laterally to the ventral premento-salivarial muscle and inserts on the anterior glossal plate. | http://purl.obolibrary.org/obo/HAO_0000377 | |
| premento-palpal muscle | The labial muscle that arises from the prementum and inserts on the first sclerite of the labial palp. | http://purl.obolibrary.org/obo/HAO_0000314 | |
| pmn-pgs | premento-paraglossal muscle | The labial muscle that arises from the ventral part of the prementum, anterior to the origin of the premento-glossal muscle and ventral premento-salivarial muscle and inserts just distally of the distal margin of the prementum. | http://purl.obolibrary.org/obo/HAO_0000687 |
| pmn | prementum | The sclerite that is median, is connected via conjunctiva along its proximolateral margins to the stipites, is articulated with the labial palps, is continuous along its distal margin with the ligula and distolateral margins with the distal hypopharynx and receives the site of attachments of the extrinsic labial palp muscles. | http://purl.obolibrary.org/obo/HAO_0000804 |
| pcs | principal carina of the stipes | The flange that extends along the margin of the posterior stipital wall. | http://purl.obolibrary.org/obo/HAO_0002099 |
| proximal galeal brush | The row of setae that is on the distal margin of the proximolateral galeal sclerite. | http://purl.obolibrary.org/obo/HAO_0002135 | |
| pgs | proximolateral galeal sclerite | The sclerite that is located on the lateral wall of the galeo-lacinial complex and bears the proximolateral galeal setiferous patch. | http://purl.obolibrary.org/obo/HAO_0002130 |
| prs | proximolateral galeal setiferous patch | The setiferous patch that is located on the lateral wall of the galeo-lacinial complex proximally of the distolateral setiferous patch. | http://purl.obolibrary.org/obo/HAO_0002129 |
| pmf | proximomedial stipital flange | The medial part of the principal carina of the stipes that is overlapped distally by the distomedial stipital carina and overlaps the lateral premental face and the proximal part of the prementum. | http://purl.obolibrary.org/obo/HAO_0002218 |
| svr | salivarium | The area that is at the proximal end of the salivary duct and corresponds to the site of insertion of the dorsal and ventral premento-salivarial muscles. | http://purl.obolibrary.org/obo/HAO_0000906 |
| svd | salivary duct | The duct that leads from the salivary gland. | http://purl.obolibrary.org/obo/HAO_0002236 |
| salivarial sclerite | The sclerite that is located in the ventral wall of the salivarium and corresponds to the site of insertion of the ventral premento-salivarial muscle. | http://purl.obolibrary.org/obo/HAO_0001682 | |
| sclerite | The area of the integument that is strongly sclerotised, with thick exocuticle and is surrounded by conjunctivae. | http://purl.obolibrary.org/obo/HAO_0000909 | |
| second intrinsic muscle of the maxillary palp | The muscle that arises from the first sclerite of the maxillary palp and inserts on the third sclerite of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0002115 | |
| second intrinsic muscle of the labial palp | The muscle that arises from the second sclerite of the labial palp and inserts on the third sclerite of the labial palp. | http://purl.obolibrary.org/obo/HAO_0002238 | |
| second sclerite of the labial palp | The sclerite that is ringlike and is connected distally to the first sclerite of the labial palp via conjunctiva and muscle. | http://purl.obolibrary.org/obo/HAO_0002195 | |
| second sclerite of the maxillary palp | The sclerite that is ringlike and is connected distolaterally to the first sclerite of the maxillary palp via conjunctiva and muscle. | http://purl.obolibrary.org/obo/HAO_0002111 | |
| seta | The sensillum that is multicellular and consists of trichogen, tormogen, and sense cells and the cuticle secreted by and adjacent to the trichogen cell. | http://purl.obolibrary.org/obo/HAO_0000935 | |
| skeletal muscle | The muscle that is attached at either end to the cuticle. | http://purl.obolibrary.org/obo/HAO_0001922 | |
| spiculate patch of galea | The area on the galeo-lacinial complex distal to the lacinial lobe that is covered with acanthae. | http://purl.obolibrary.org/obo/HAO_0002139 | |
| spiculate patch of the lacinia | The area on the lacinial lobe that is covered with acanthae. | http://purl.obolibrary.org/obo/HAO_0002138 | |
| spine | The process that lacks non-sclerotised ring at the base. | http://purl.obolibrary.org/obo/HAO_0000949 | |
| stipes, stipites | The appendage that is connected posteroproximally to the hypostoma, anteroproximally and lateroproximally to the mandible and medioproximally to the labium and the hypopharynx via conjunctiva, is connected to the cranium via muscles and that bears the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0000958 | |
| stipital sclerite | The sclerite that is on the medial stipital wall, bears the medial stipital process. | http://purl.obolibrary.org/obo/HAO_0002096 | |
| sti-gal | stipito-galeal muscle | The maxillar muscle that arises from the posterior sclerite of the stipes medial to the site of origin of the stipito-lacinial muscle and inserts on the median wall of the galeo-lacinial complex distal to the stipito-lacinial muscle. | http://purl.obolibrary.org/obo/HAO_0001661 |
| sti-lac | stipito-lacinial muscle | The maxillar muscle that arises along the lateral margin of the posterior stipital sclerite and inserts proximally on the median wall of the stipes just proximal to the lacinal lobe. | http://purl.obolibrary.org/obo/HAO_0001660 |
| sti-mp1 | stipito-palpal muscle | The maxillary muscle that arises from the posterior stipital sclerite and inserts on the first sclerite of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0002110 |
| stipito-premental conjunctiva | The conjunctiva that extends along the posterior (dorsal) margin of the premental arm and the proximal margin of the medial stipital wall. | http://purl.obolibrary.org/obo/HAO_0002125 | |
| sch | stipitocardinal hinge | The membranous area linking the cardo and stipes. | http://purl.obolibrary.org/obo/HAO_0002076 |
| sts | styloconic sensillum | The seta that is on a process. | http://purl.obolibrary.org/obo/HAO_0002211 |
| tnt-cd | tentorio-cardinal muscle | The maxillary muscle that arises from the anterior region of the cranium and inserts adjacent to the cardino-stipital hinge. | http://purl.obolibrary.org/obo/HAO_0001638 |
| tnt-sti | tentorio-stipital muscle | The maxillar muscle that arises on the posteroventral part of the anterior tentorial arm and inserts on the median wall of the stipes. | http://purl.obolibrary.org/obo/HAO_0001002 |
| tentorium | The apodeme that has its sites of origin marked by the anterior and posterior tentorial pits and gular sulci. | http://purl.obolibrary.org/obo/HAO_0001003 | |
| third intrinsic muscle of the maxillary palp | The muscle that arises from the second sclerite of the maxillary palp and inserts on the third sclerite of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0002116 | |
| third sclerite of the labial palp | The sclerite that is ringlike and is connected distally to the second sclerite of the labial palp via conjunctiva and muscle. | http://purl.obolibrary.org/obo/HAO_0002196 | |
| 3mp | third sclerite of the maxillary palp | The sclerite that is ringlike and is connected distally to the second segment of the maxillary palp via conjunctiva and muscle. | http://purl.obolibrary.org/obo/HAO_0002112 |
| ss1 | uniporous sensilla, Type 1 seta | The sensillum whose cuticular component has one cuticular pore. | http://purl.obolibrary.org/obo/HAO_0002221 |
| vlm | velum | The flange that is transparent, and extends along the anterodistal margin of the galeo-lacinial complex distal to the lacinial lobe. | http://purl.obolibrary.org/obo/HAO_0002140 |
| vdp | ventral dististipital process | The projection that is located distally on the posterior stipital sclerite and encircles the base of the maxillary palp. | http://purl.obolibrary.org/obo/HAO_0002102 |
| ventral glossal lines | The carinae that radiates from the posterior glossal plate towards the apical glossal setae. | http://purl.obolibrary.org/obo/HAO_0002230 | |
| vpf | ventral premental face | The area of the prementum that is delimited laterally by the lateral premental face. | http://purl.obolibrary.org/obo/HAO_0002156 |
| pmnv-slv | ventral premento-salivary sclerite muscle | The salivarial muscle that arises from the proximal end of the ventral part of the prementum and inserts on the salivarial sclerite. | http://purl.obolibrary.org/obo/HAO_0001072 |
| vba | ventrolateral basiglossal arm | The projection that is located proximolaterally on the anterior glossal plate and articulates with the basiparaglossal sclerite. | http://purl.obolibrary.org/obo/HAO_0002202 |
Volume rendered CLSM media files on figshare.com
https://doi.org/10.6084/m9.figshare.861056
https://doi.org/10.6084/m9.figshare.861057
https://doi.org/10.6084/m9.figshare.861058
https://doi.org/10.6084/m9.figshare.861060
https://doi.org/10.6084/m9.figshare.861061
https://doi.org/10.6084/m9.figshare.861062
https://doi.org/10.6084/m9.figshare.861063
https://doi.org/10.6084/m9.figshare.861064
https://doi.org/10.6084/m9.figshare.861065
https://doi.org/10.6084/m9.figshare.861066
https://doi.org/10.6084/m9.figshare.861067
https://doi.org/10.6084/m9.figshare.861119
https://doi.org/10.6084/m9.figshare.956282
https://doi.org/10.6084/m9.figshare.956281
https://doi.org/10.6084/m9.figshare.956280