Research Article |
|
Corresponding author: Alexey Reshchikov ( lerth@yandex.ru ) Corresponding author: Jing-Xian Liu ( liujingxian@yahoo.com ) Academic editor: Gavin Broad
© 2021 Xin-Fang Zheng, Alexey Reshchikov, Jing-Xian Liu.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Zheng X-F, Reshchikov A, Liu J-X (2021) Two new species of Drepanoctonus Pfankuch, 1911 (Hymenoptera, Ichneumonidae) from the Oriental region. Journal of Hymenoptera Research 83: 1-19. https://doi.org/10.3897/jhr.83.66400
|
Abstract
Two new species of the genus Drepanoctonus Pfankuch, 1911 collected in natural habitats with Wild Tea (Camellia sinensis var. assamica) are described and illustrated: D. rimdahli Liu & Reshchikov, sp. nov. from Chiang Mai Province, Thailand and D. chamagudao Liu & Zheng, sp. nov. from Yunnan Province, China. Drepanoctonus bicolor Kusigemati, 1971 is recorded from China for the first time. An identification key to the species of the genus is provided.
Keywords
Camellia sinensis assamica, Darwin wasps, Metopiinae, new species, parasitoids, Tea Fauna, Wild Tea
Introduction
Drepanoctonus Pfankuch, 1911 is a small genus in the subfamily Metopiinae (Hymenoptera, Ichneumonidae), comprising six known species. Three species are known from the Palaearctic region (D. bicolor Kusigemati, 1971, D. tibialis Pfankuch, 1911 and D. tricoloratus (Šedivý, 1971)), one from the Oriental region (D. auritus Chiu, 1962), one from the Afrotropical region (D. bicinctus (Benoit, 1961)), and one from the Australian region (D. bifasciatus (Brullé, 1846)) (
In the present study, two species are described as new to science: Drepanoctonus chamagudao Liu & Zheng, sp. nov. from Southwest China, and Drepanoctonus rimdahli Liu & Reshchikov, sp. nov. from Northern Thailand (Fig.
Materials and methods
The specimens examined are deposited in the following institutions (curators in parenthesis):
The specimens of Drepanoctonus rimdahli Liu & Reshchikov, sp. nov. were collected in Northern Thailand by Malaise trap during the “Tea Fauna” project (http://teafauna.com) in the understory of an old secondary forest with Camellia sinensis var. assamica (Masters) Kitamura (Fig.
Specimens were examined using the Zeiss Stemi 508 stereomicroscope. Images were acquired digitally using the KEYENCE VHX-5000 Digital Microscope Imaging System, Leica S8APO Digital Microscope System and processed with Adobe Photoshop.
Morphological terminology and nomenclature of wing venation follows
Molecular analysis
Genomic DNA of the new species was extracted from two females and one male using DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany), following a non-destructive DNA extraction protocol as described in
Results
Following the non-destructive extraction of DNA, the three specimens were identified as two new species: Drepanoctonus chamagudao sp. nov. (MW528531) and Drepanoctonus rimdahli sp. nov. (MW528532; MW528533; MW528534), as described below. These two distinct species are also supported by the COI sequences, pairwise percentage identify of the sequences was 84.4%–84.6% (interspecies distance between D. chamagudao sp. nov. and D. rimdahli sp. nov), and 99.9%–100% (intraspecies distance of D. rimdahli sp. nov.).
Taxonomy
Order Hymenoptera
Family Ichneumonidae
Subfamily Metopiinae
Drepanoctonus
Drepanoctonus
Pfankuch, 1911: 688. Type species: Drepanoctonus tibialis Pfankuch. Designated by
Zonopius
Benoit, 1961, 63: 305. Type species: Zonopius bicinctus Benoit. Original designation. Synonymized by
Generic diagnosis
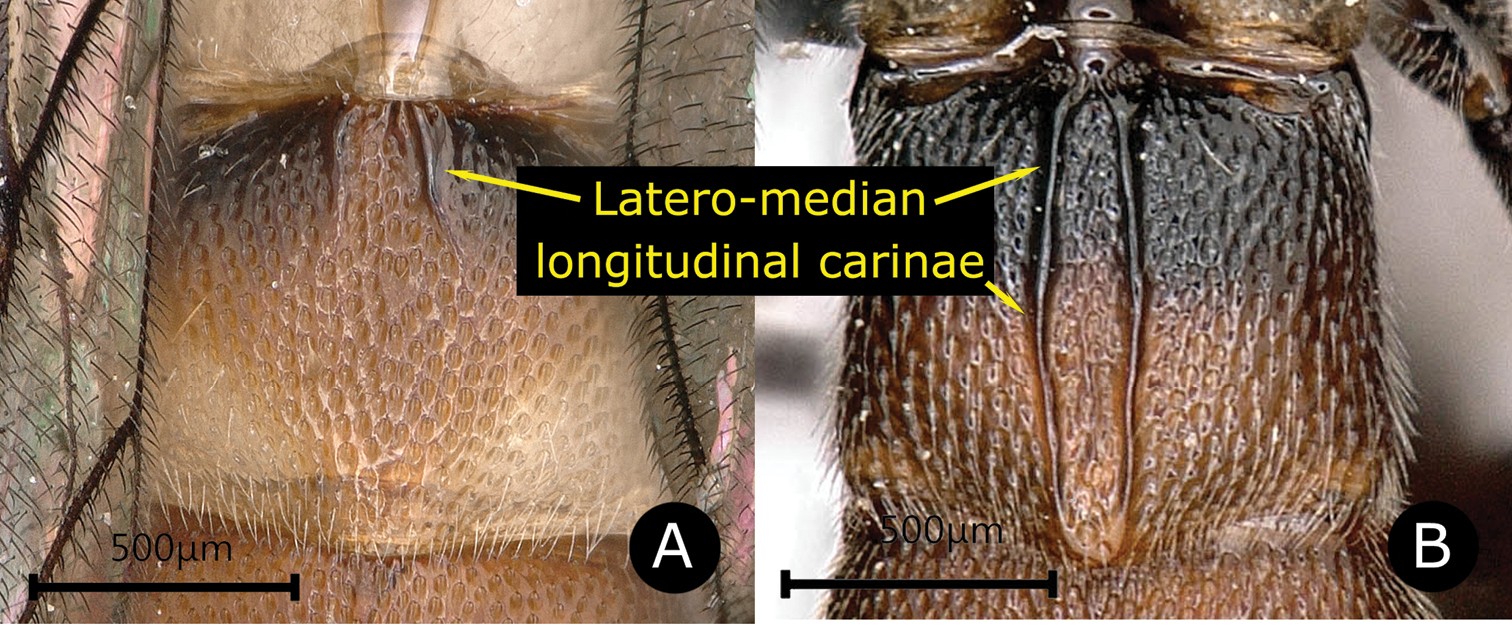
Fore wing length 6.0–9.0 mm. Body with punctures rather sharp and dense. Combined face and clypeus weakly convex; upper margin of face produced medially as an acute triangle between bases of antennae (except D. rimdahli sp. nov.). Pronotum posteriorly with a swelling just below its upper margin. Epicnemial carina with upper end far from the front edge of mesopleuron. Mesopleuron moderately convex. Propodeum rather short, latero-median longitudinal carinae complete, anterior transverse carina absent, area superomedia confluent with area basalis; posterior transverse carina complete or interrupted in the middle. Propodeal spiracle elongate. Spurs of middle tibia elongate, approximately equal in length. Fore wing with 1cu-a opposite or distad to M&RS, and 2rs-m nearly opposite to 2m-cu. T1 with an oblique baso-dorsal edge, with latero-median longitudinal carinae strong and sharp to apex. T2 usually with a pair of latero-median longitudinal carinae, either shortly present on base or reaching to posterior margin of tergite. T3 and T4 with or without a single weak, incomplete median longitudinal carina. Laterotergite of T2 vestigial, that of T3 narrowly wedge shaped, and that of T4 to T6 moderately wide and separated from their tergites by a crease (
Distribution
Palaearctic, Oriental, Australian and Afrotropical regions (Fig.

A distribution of the genus Drepanoctonus Pfankuch, 1911 B type habitat of D. rimdahli Liu & Reshchikov, sp. nov. C D. tibialis Pfankuch, 1911 with its host, Watsonalla binaria (Hufnagel, 1767), Germany, Saxony, Radeburg-Berbisdorf, 18 Oct. 2016, ex. 25 May 2017, Franziska Bauer leg. (pers. comm. Bauer)
Biology
Parasitoids of Drepanidae (Lepidoptera) (
Key to species of Drepanoctonus (modified from Tolkanitz, 1987)
| 1 | Latero-median longitudinal carinae of T2 either absent, only present on anterior 0.3 of the tergite, or present as foveolate lines (Fig. A) | 2 |
| – | Latero-median longitudinal carinae of T2 strong and reaching the posterior margin of the tergite (Fig. B) | 4 |
|
||
| 2 | T2 medially with a pair of foveolate lines at base; T3 to T5 medially with a depressed triangular area at base; fore wing with 1cu-a oblique and almost opposite to M&RS; hind tibia entirely reddish brown without white or yellow band; upper margin of pronotum yellow; T4 with a yellow band posteriorly (Fig. A) | D. bicinctus (Benoit, 1961) |
| – | T2 medially either without foveolate lines or carinae, or anteriorly with a pair of short latero-median longitudinal carinae; T3 to T5 without depressed triangular area at base; fore wing with 1cu-a distinctly distad of M&RS; hind tibia reddish brown with a sub-basal white or yellow band; upper margin of pronotum reddish brown or entirely black; T4 without a yellow band (Fig. B) | 3 |

|
||
| 3 | Upper margin of face not produced backward as inter-antennal projection; area superomedia of propodeum polished and glabrous; T2 with a pair of short latero-median longitudinal carinae on anterior 0.3 of tergite (Fig. A); T3 evenly reddish brown | D. rimdahli sp. nov. |
| – | Upper margin of face produced backward forming an inter-antennal projection; area superomedia of propodeum with several transverse wrinkles; T2 without a pair of short latero-median longitudinal carinae (Fig. B); T3 posteriorly with a wide yellow band | D. bifasciatus (Brullé, 1846) |

|
||
| 4 | T3 without latero-median longitudinal carinae (Fig. A) | 5 |
| – | T3 with latero-median longitudinal carinae (Fig. B) | 6 |

|
||
| 5 | T2 to T6 transverse, roughly wrinkled and punctate; metasoma black, T1 to T6 with their apical margins reddish–brown; femora black with apex yellow; all tibiae yellow | D. tibialis Pfankuch, 1911 |
| – | T2 to T6 not transverse, punctate; metasoma with T1 and T2 black, T3 to T6 reddish; femora entirely black, fore tibia reddish-brown, mid and hind tibia black | D. bicolor Kusigemati, 1971 |
| 6 | Posterior transverse carina of propodeum complete (Fig. A); fore wing with 1cu-a opposite to M&RS; T3 with a pair of latero-median longitudinal carinae on anterior half (Fig. A), T4 without carina; T1–T3 usually red, T5 posteriorly with round white spot | D. tricolorata (Šedivý, 1971) |
| – | Posterior transverse carina of propodeum incomplete, dorso-medially absent (Fig. B); fore-wing with 1cu-a distinctly distad of M&RS; both T3 and T4 with a single median longitudinal carina (Fig. B) | 7 |

|
||
| 7 | Metasoma mainly blackish–brown, T1 with a pair of triangular yellow spots on latero-posterior corners; T2 with a yellow band on posterior 0.2–0.4 which is interrupted by latero-median longitudinal carinae (Fig. A) | D. auritus Chiu, 1962 |
| – | Metasoma mainly reddish, T1 black with a pair of yellow spots on latero-posterior corners, T2 with anterior half black and posterior half reddish (Fig. B) | D. chamagudao sp. nov. |

|
Drepanoctonus bicolor
Drepanoctonus bicolor Kusigemati, 1971, 250.
Materials examined
China: 1 male, Shanxi province, Mt. Lishan, Xiahe Protection Area, 35°20.40'N, 112°32.41'E, 780 m, 22–25 Aug. 2012, Ren Ya-Jun leg. (
Comments

Distribution
China (Shanxi Province), Russia, Japan.
Drepanoctonus chamagudao , sp. nov.
Materials examined
Holotype, female. China: Yunnan Province, Yuxi City, Xinping County, Mt. Ailao, Cha Ma Gu Dao, 98°53'43.38"N, 28°18'56.4984"E, 2538 m, 8. Aug. 2018, Zheng Xin-Fang leg., DNA voucher,
Descriptions
Female. Fore wing length 7.5 mm, body length 8.0 mm (Fig.



Head. Combined face and clypeus densely and strongly punctate, densely setose, 0.7× as wide as high (Fig.
Mesosoma. Pronotum strongly punctate and setose on upper half, more or less shiny and glabrous on lower half, with a row of short and transverse wrinkles along posterior margin. Epomia strong, reaching upper 0.8 of pronotum. Mesoscutum strongly punctate and setose (Fig.
Wings. Fore wing with 1cu-a distad of M&RS, separated from M&RS by 0.67× its own length, 2m-cu almost opposite to 2rs-m, 3rs-m absent. Hind wing with Cu & cu-a interrupted above the middle, distal abscissa of Cu weakly pigmented.
Legs. Fore and mid claws with 1–2 pectinate teeth at base. Mid tibial spurs nearly equal in length. Hind femur 4.8× as long as its maximum width, hind tibia 5.8× as long as its maximum width, hind tibial spurs equal in length, 0.57× as long as 1st segment of tarsus (Fig.
Metasoma. T1 1.0× as long as its apical width, punctate, latero-median longitudinal carinae strong, anterior base of T1 strongly oblique in lateral view, dorsolateral carina sharp and complete, spiracle located on anterior 0.3 of the tergite (Fig.
Colour. Head and mesosoma black, covered with whitish setae (Fig.
Male. Unknown.
Distribution
China (Yunnan province).
Comments
This species is similar to D. bicolor (Fig.
It also very resembles Drepanoctonus auritus Chiu, 1962 (Fig.
Etymology
The species is named after the type locality, Chamagudao, which means the Ancient Tea Horse Road of Yunnan province.
Drepanoctonus rimdahli , sp. nov.
Materials examined
Holotype, female, Thailand: Chiang Mai, Mae Taeng, Pa Pae, 19°14'30.6"N, 98°30'14.1"E, old forest with C. sinensis assamica, Malaise trap (Dara#1), 04.V-25.V.2017, Monsoon Tea leg. (
Description
Holotype. Female, fore wing length 7.5 mm, body length 9.0 mm (Fig.

Head. Combined face and clypeus weakly convex, 0.80–0.90× as wide as high, densely and evenly punctate, lateral corner of clypeus weakly wrinkled (Fig.

Mesosoma. Pronotum with upper lateral corner finely punctate and densely setose, lower part polished with a row of transverse depressions. Epomia strong, reaching to upper 0.8 of pronotum. Mesoscutum with minute punctures, distance between punctures 0.5–1.0× the diameter of a puncture (Fig.
Wings. Fore wing with 1cu-a distad of M&Rs by 0.5× the length of 1cu-a, 2rs-m slightly reclivous, very slightly anterior of 2m-cu. Hind wing with Cu & cu-a interrupted on lower 0.4, distal abscissa of Cu distinct.
Legs. Mid tibial spurs equal in length and 0.63× the length of 1st segment of hind tarsus. Hind femur 4.0× as long as its maximum width (Fig.
Metasoma. T1 1.25× as long as its posterior width, latero-median longitudinal carinae weak but forming a longitudinal convex area, lateral area of T1 subpolished, with sparse minute punctures, dorsolateral carina complete and sharp (Fig.
Colour. Body with whitish setae. Head black (Fig.
Male. (Fig.
Variation
T2 of a female paratype, with a pair of short carinae on anterior 0.4 of tergite. Pronotum largely black with upper margin tinged with indistinct dark reddish-brown. Mesopleuron weakly marked with dark reddish-brown.
Distribution
Thailand (Chiang Mai).
Comments
This species can be separated from other species of the genus by the following combined characters: the upper margin of face not produced medially as an acute triangle between bases of antennae (Fig.
Etymology
The species is named after Mr. Kenneth Rimdahl, the founder of Monsoon Tea, in recognition of his efforts in saving Thai forests.
Discussion
In most species of Drepanoctonus the upper margin of the face is distinctly produced medially as an acute triangle between bases of antennae, and the upper side of the triangle is deeply grooved. This character is absent in D. rimdahli sp. nov., but we place this species in the genus Drepanoctonus because all other diagnostic characters match the generic description. Two species, D. rimdahli sp. nov. and D. bicinctus, have a pair of short stubs or incomplete latero-median longitudinal carinae on their second metasomal tergite (Fig.
Because of the lack of fresh material for the known species of Drepanoctonus, only the COI sequences of the two new species is available as an additional evidence for identification and future molecular analysis.
The scattered worldwide distribution of the genus Drepanoctonus (Fig.
Acknowledgements
The authors are deeply grateful to the team of Monsoon Tea and its founder Kenneth Rimdahl for assistance in sampling in Northern Thailand; William Persson for type locality photography, Wang Peng for his kind help in field work in Yunnan; Ren Ya-Jun (South China Agricultural University) for collecting the specimens from Shanxi; Chi-Feng Lee (
References
- Aubert JF (1965) Les Ichneumonides du rivage méditerranéen français (8e serie, Region Cotiere entre La Ciotat et Saint-Tropez). Vie et Milieu 16: 549–573.
- Benoit PLG (1961) Nouveaux Metopiinae de Madagascar (Hym. Ichneumonidae). Revue de Zoologie et de Botanique Africaines 63: 299–308.
- Broad GR, Shaw MR, Fitton MG (2018) Ichneumonid wasps (Hymenoptera: Ichneumonidae): their classification and biology. Handbooks for the Identification of British Insects 7(12): 1–418.
- Capek M, Hladil J, Šedivý J (1982) Verzeichnis der aus verschiedenen Insekten erzogenen parasitschen Hymenopteren – Teil VI. Entomological Problems (Bratislava) 17: 325–371.
- Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3(5): 294–299.
- GBIF Secretariat (2019) GBIF Backbone Taxonomy. Checklist dataset. https://doi.org/10.15468/39omei [accessed via https://www.gbif.org/species/1285027 on 2020-08-27]
- Horstmann K (1986) Typenrevision der von Karl Pfankuch beschriebenen Arten und Formen der Familie Ichneumonidae (Hymenoptera). Entomologische Mitteilungen aus dem Zoologischen Museum Hamburg 8: 251–264.
- Kusigemati K (1971) Taxonomic studies on the subfamily Metopiinae of Japan (Hymenoptera: Ichneumonidae). Memoirs of the Faculty of Agriculture, Kagoshima University 8(1): 205–298.
- Pfankuch K (1911) Die Ichneumonidengattung Drepanoctonus Kriechb. (Hym.). Deutsche Entomologische Zeitschrift 1911: 687–689. https://doi.org/10.1002/mmnd.48019110613
- Pfankuch K (1935) Verzeichnis der Ichneumoniden von Bremen und Umgegend. Mitteilungen aus dem Entomologischen Verein in Bremen 22(1934): 6–30.
- Reshchikov A, Santos BF, Liu JX, Barthélémy C (2019) Review of Palpostilpnus Aubert (Hymenoptera, Ichneumonidae, Phygadeuontinae), with the description of ten new species. European Journal of Taxonomy 582: 1–63. https://doi.org/10.5852/ejt.2019.582
- Taekul C, Valerio AA, Austin AD, Klompen H, Johnson NF (2014) Molecular phylogeny of telenmine egg parasitoids (Hymenoptera: Platygastridae s.l: Telenominae): evolution of host shifts and implications for classification. Systematic Entomology 39: 24–35. https://doi.org/10.1111/syen.12032
- Tolkanitz VI (1987) Parasitic Hymenoptera. Ichneumonidae- Metopiinae. Fauna Ukraina 11(2): 1–212. [In Russian]
- Townes HK (1971) The genera of Ichneumonidae, Part 4. Memoirs of the American Entomological Institute 17: 1–372.
- Yu DS, van Achterberg C, Horstmann K (2016) Taxapad 2016, Ichneumonoidea. Database on flash-drive, Nepean.
Supplementary material
fasta file of COI sequences of some species
Data type: sequence of COI
Explanation note: COI sequences of different individual speciemns from the two new species.