Research Article |
|
Corresponding author: Simon van Noort ( svannoort@iziko.org.za ) Academic editor: Gavin Broad
© 2021 Simon van Noort, Zachary Lahey, Elijah J. Talamas, Andrew D. Austin, Lubomir Masner, Andrew Polaszek, Norman F. Johnson.
This is an open access article distributed under the terms of the CC0 Public Domain Dedication.
Citation:
van Noort S, Lahey Z, Talamas EJ, Austin AD, Masner L, Polaszek A, Johnson NF (2021) Review of Afrotropical sceliotracheline parasitoid wasps (Hymenoptera, Platygastridae). In: Lahey Z, Talamas E (Eds) Advances in the Systematics of Platygastroidea III. Journal of Hymenoptera Research 87: 115-222. https://doi.org/10.3897/jhr.87.73770
|
Abstract
The platygastrid subfamily Sceliotrachelinae, while represented globally by numerous biologically important taxa, is generally poorly known for the Afrotropical region. It contains a number of species of economic significance, including some that attack hemipteran pests of citrus. Here we review the taxonomy of the subfamily for the region. We revise two enigmatic, endemic South African genera, Afrisolia Masner and Huggert and Sceliotrachelus Brues, providing illustrated identification keys to the species. Afrisolia anyskop van Noort & Lahey, sp. nov., Afrisolia quagga van Noort & Lahey, sp. nov. and Afrisolia robertsoni van Noort & Lahey, sp. nov. are described for this previously monotypic genus. Sceliotrachelus was previously known only from the two male type specimens of Sceliotrachelus braunsi Brues. Two new species, Sceliotrachelus midgleyi van Noort, sp. nov. and Sceliotrachelus karooensis van Noort, sp. nov., are described from the Eastern and Western Cape, respectively. The female of S. braunsi is described for the first time and additional distribution records for the species are documented. The putatively basal species, S. karooensis, exhibits less derived morphology than the highly apomorphic S. braunsi, necessitating reassessment of the limits of the genus. The exceptional morphology exhibited by species of Sceliotrachelus is hypothesized to be an adaptation to living in leaf-litter. We also revise the Old World species of Parabaeus Kieffer, describing a remarkable new species, Parabaeus nasutus van Noort, sp. nov., and provide an illustrated and updated key to the species. Fidiobia Ashmead is currently represented in Africa by seven described species to which we add Fidiobia celeritas van Noort & Lahey, sp. nov., a charismatic new species from South Africa. The distribution of the genus Isolia is expanded to include new country records for I. hispanica Buhl (Kenya) and an undescribed species from Madagascar. To facilitate the ongoing exploration and documentation of the African platygastrid fauna, an identification key to Afrotropical sceliotracheline genera and high resolution images of exemplar species for the remaining genera are provided. An overview of known species richness and biology is also included. All images presented here as well as additional images and interactive online Lucid identification keys are available on WaspWeb at http://www.waspweb.org.
Keywords
Adaptation, Africa, holotype, identification key, morphology, new species, taxonomy
Introduction
The subfamily Sceliotrachelinae was erected by
The Afrotropical sceliotracheline fauna is currently represented by 35 described species (number listed in brackets following each genus) and numerous undescribed species contained in eight genera: Afrisolia Masner & Huggert, 1989 (1 species); Allotropa Förster, 1856 (7 species); Amitus Haldeman, 1850 (only undescribed species); Fidiobia Ashmead, 1894 (7 species); Isolia Förster, 1878 (1 species); Parabaeus Kieffer, 1910 (8 species); Pulchrisolia Szabó, 1959 (10 species); and Sceliotrachelus Brues, 1908 (1 species) (
We review the Afrotropical Sceliotrachelinae, revise two enigmatic, endemic South African genera (Afrisolia and Sceliotrachelus), revise the Old World species of Parabaeus, and provide illustrated identification keys to the species in these genera. To facilitate the ongoing exploration and documentation of the African fauna, identification keys to the Afrotropical sceliotracheline genera are provided, as well as high resolution images of exemplar species for each genus, including an overview of known species richness and biology. All images presented here, as well as additional images and interactive online identification keys are available on WaspWeb at http://www.waspweb.org.
Materials and methods
Images were acquired at SAMC with a Leica LAS 4.9 imaging system, comprising a Leica Z16 microscope (using either a 2× or 5× objective) with a Leica DFC450 Camera and 0.63× video objective attached. The imaging process, using an automated Z-stepper, was managed using the Leica Application Suite V 4.9 software installed on a desktop computer. Diffused lighting was achieved using a Leica LED5000 HDI dome. Scanning Electron Micrographs were acquired using a Phenom XL G2 Desktop SEM at 5 keV on uncoated specimens. Specimens from NHMUK were imaged using a Canon DSLR with 10× Mitutoyo objective and processed with Helicon Focus stacking software. Post-processing of photographs and line drawings were done using Adobe Photoshop 2021. Plate production for figures was done using Adobe Illustrator 2021. All images presented in this paper, as well as supplementary images, are available on WaspWeb at www.waspweb.org.
Morphological terminology follows
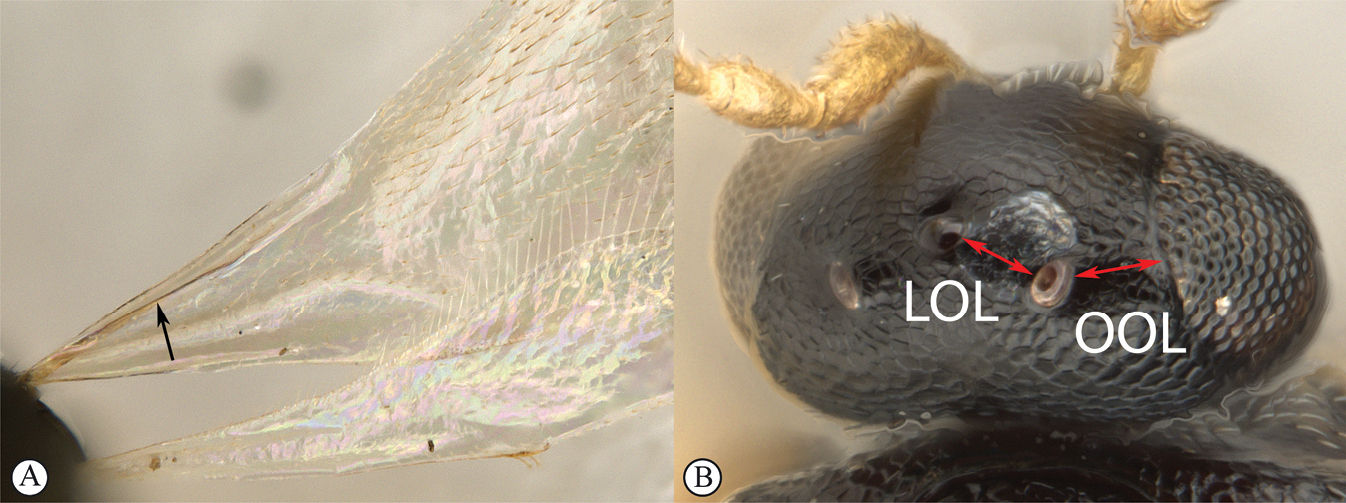
LOL lateral ocellar line, shortest distance between inner margins of median and lateral ocelli
OOL ocular ocellar line, shortest distance from inner orbit and outer margin of posterior ocellus
POL posterior ocellar line, shortest distance between inner margins of posterior ocelli
Delimitation of the Afrotropical biogeographical region is based on the concept of the old Ethiopian region of
Depositories
MZLU Lund Museum of Zoology, Lund, Sweden
OSUC C.A. Triplehorn Insect Collection, Columbus, Ohio
SANC National Collection of Insects, Pretoria
WARI Waite Agricultural Research Institute, Adelaide
Accession/catalogue numbers prefixed with OSUC are unique identifiers for individual specimens, whereas numbers prefixed with SAM-HYM are unique identifiers at either the specimen level, or as a series of conspecific specimens with the same collecting data. These may be individually circumscribed by the addition of an alphabetic suffix.
Results
Sceliotrachelinae
Sceliotrachelinae
Brues, 1908: 3, 12 (original description, keyed);
Amitini
Szabó, 1959: 390 (original description. Synonymized by
Sceliotrachelini
Brues, 1908:
Diagnosis
The subfamily is poorly defined with no confirmed synapomorphic characters uniting the currently included taxa. Many, but not all, sceliotracheline genera possess foamy structures, which, among platygastroids, are limited to Platygastridae (
Key to Afrotropical genera of Sceliotrachelinae
| 1 | Wings absent (A); T1 fused with T2 and S1 fused with S2, with no evident sulcus (A, B); frons with a transverse furrow above torulus | Parabaeus |

|
||
| – | Wings fully developed (a) extending to at least T2 (b); T1 separated from T2 and S1 from S2 by a sulcus (b), sulcus sometimes partly obscured by dense pilosity (b); frons above torulus without transverse furrow | 2 |

|
||
| 2 | Spur of fore tibia combed (pts) (Isolia species-group) | 3 |

|
||
| – | Spur of fore tibia without comb | 6 |

|
||
| 3 | Pronotum with medial longitudinal sulcus (pms) (A); sternite 2 with long setae (B) | Sceliotrachelus |

|
||
| – | Pronotum without medial sulcus (a); S2 glabrous (b) | 4 |

|
||
| 4 | Fore wings and microtrichia distinctly bicolored, giving the appearance of patches or stripes | Pulchrisolia |

|
||
| – | Fore wings and microtrichia not bicolored | 5 |

|
||
| 5 | Fore wing with tubular submarginal vein (A); admedian depressions (adm) present, may be weak (A); transepisternal line present (tel) (B) | Afrisolia |

|
||
| – | Fore wing without veins (a); admedian depressions absent (a); transepisternal line absent (b) | Isolia |
|
||
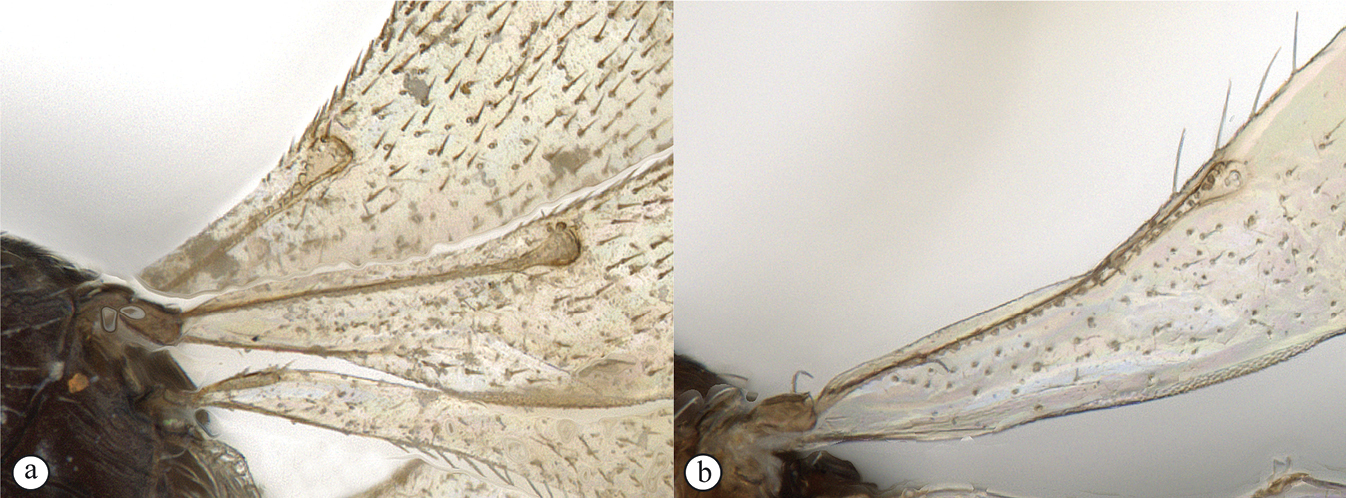
| 6 | Fore wing with rudimentary submarginal vein, only sclerotized for about half its length (A); OOL usually subequal to or longer than LOL (B); body usually short and squat | Amitus |
|
||
| – | Fore wing with submarginal vein fully sclerotized, ending in knob (a, b); OOL always distinctly shorter than LOL; body usually elongate, spindle-like | 7 |
|
||
| 7 | Propodeum medially with single keel (A); metasomal depression not visible; notauli absent (A); scutoscutellar sulcus foveolate (A); anterior T2 with a transverse line of foveae; in females, ventral A8–A9 projecting distally; male antenna with long setae; submarginal vein of fore wing distinctly surpassing basal 0.33 of wing (B) | Allotropa |

|
||
| – | Propodeum with H-shaped foamy structure (a); metasomal depression delimited by lateral propodeal carinae; notauli usually abbreviated, triangular (a), absent in a few species; scutoscutellar sulcus simple (a); anterior T2 with two pits, without a line of foveae; antennae variable, but not as above; submarginal vein of fore wing reaching at most to basal 0.33 of wing (b) | Fidiobia |
|
||
Afrisolia
Afrisolia
Masner & Huggert, 1989: 34 (original description. Type: Afrisolia obesa Masner and Huggert, by original designation);
Diagnosis
Afrisolia is the only genus in the Isolia-cluster with a transepisternal line, which is strong and in a posteriorly facing cup–shape; fore wing with short, strong tubular submarginal vein; fore wing broad, with microtrichiae on disc transformed into bulbous spiculae, marginal cilia absent; anterior admedian depressions present, but may be weak. Species are stout and squat with black head and mesosoma and brown metasoma; surface sculpture micro-alutaceous, polished, matte, or delicately reticulate; notauli at same level with mesoscutum, surrounded by deep grooves; median foamy keels of propodeum spaced widely apart, with 3 slight elevations or abbreviated costae between them.
Species richness
Afrisolia anyskop van Noort & Lahey, sp. nov. (South Africa) (Figs
Afrisolia obesa Masner & Huggert, 1989 (South Africa) (Fig.
Afrisolia quagga van Noort & Lahey, sp. nov. (South Africa) (Figs
Afrisolia robertsoni van Noort & Lahey, sp. nov. (South Africa) (Fig.
Distribution
Afrotropical: South Africa (
Biology
Unknown.
Comments
The Afrisolia species recorded from Iran (represented by two males) in
Key to species of Afrisolia
| 1 | Gena distinctly concave in anterior view (A); pronotal shoulders well-developed, pronotum anteriorly with well-defined neck (B); notauli narrow, 7× longer than wide (B) | Afrisolia anyskop van Noort & Lahey, sp. nov. |

|
||
| – | Gena straight or convex in anterior view (a); pronotal shoulders more evenly rounded, pronotum anteriorly with weakly defined neck (b); notauli broad, at most 5× longer than wide (b) | 2 |

|
||
| 2 | Metasoma dark brown, not strongly contrasting with head and mesosomal coloration (A); notauli 5× longer than wide (B); OOL equal to LOL (C) | Afrisolia robertsoni van Noort & Lahey, sp. nov. |

|
||
| – | Metasoma yellow-orange or red-orange, contrasting with darker head and mesosoma (a); notauli at most 3× longer than wide (b); OOL either distinctly shorter (c) or distinctly longer than LOL | 3 |

|
||
| 3 | Notauli posteriorly widely spaced relative to scutellar width (red lines), meeting the mesoscutellar sulcus less than the posterior notaular width from the lateral margins of the scutellum (white lines) (A); mesoscutum polished between the notauli (A); OOL shorter than LOL, two-fifths of POL, (B); single dorsal striation on mesopleuron (C) | Afrisolia obesa Masner & Huggert |

|
||
| – | Notauli positioned closer together relative to scutellar width (red lines), meeting the mesoscutellar sulcus at least double the posterior notaular width from the lateral margins of the scutellum (white lines) (a); mesoscutum with alutaceous reticulation between the notauli (a); OOL longer than LOL, two-thirds of POL (b); two or more dorsal striations on mesopleuron (c) | Afrisolia quagga van Noort & Lahey, sp. nov. |

|
||
Afrisolia anyskop , sp. nov.
Material examined
Holotype
: South Africa • ♀; Western Cape, Anyskop Farm, (5.5 km 290°W Langebaanweg); 32°57.301'S, 18°05.283'E; 18–25 September 2002; S. van Noort; Yellow pan; LW02-N8-Y282; Sand Plain Fynbos; SAM-HYM-P032464 (
Paratypes
: South Africa • 1 ♂; data as for holotype except for LW02-N8-Y288, SAM-HYM-P032465 (
Description
Female. Body length 1.05 mm. Colour of body black, with metasoma, antennae, alar sclerites, coxae, fore femur, and fore tibia dark brown, meso- and meta-tibiae, all tarsi, and mandibles yellow-brown; wings appear white due to dense presence of white microtrichiae.

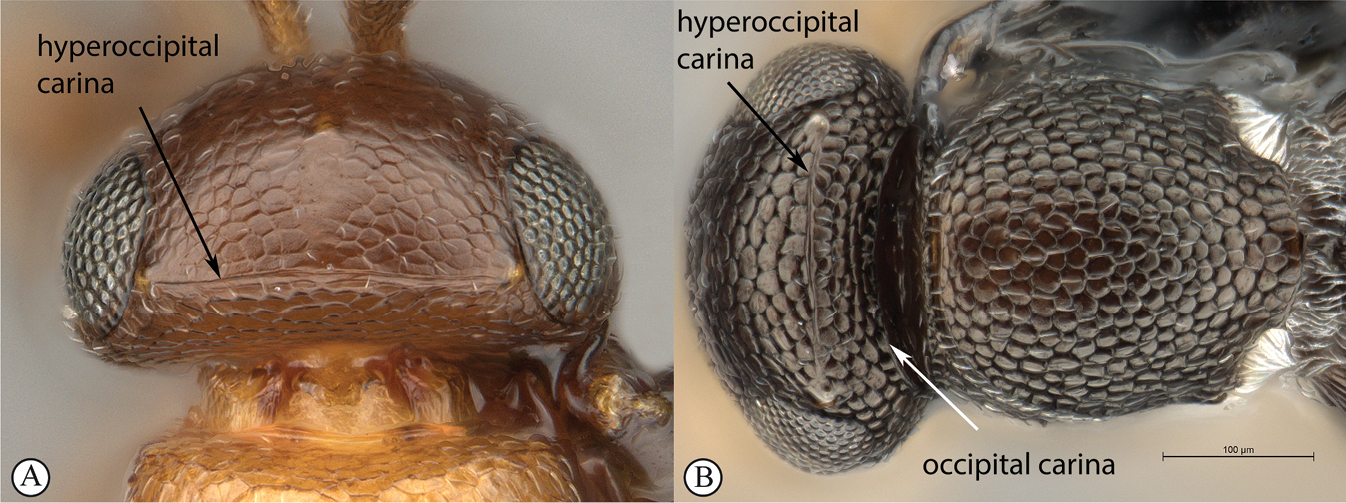
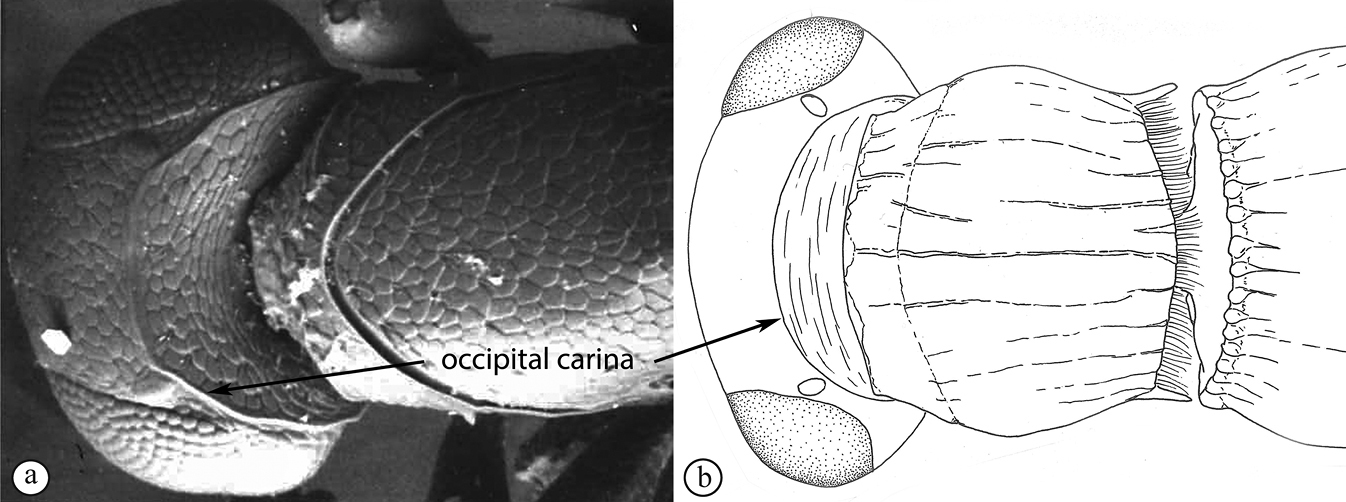
Head in dorsal view strongly transverse (7:24), lens-like, with frons only gently arched; temples straight, strongly receding toward occipital carina; POL:LOL:OOL = 7:3:3. Scape distinctly shorter than interorbital space (5:9) with scrobe absent; eyes with minute hairs, scattered setae on head slightly longer; head with delicate alutaceous reticulation, including posterior of hyperoccipital carina; head in lateral view almost twice as high as wide (5:9), with lower frons gently convex, vertex acute and posterior toruli distinctly raised; eyes oval (13:23) and upper part of gena almost disappearing behind eye; posterior edge of gena defined by crenulate and sharp occipital carina with c. 5 long, posteriorly projecting setae from ventral section of carina; malar space almost as long as eye height (45:55); head in anterior view wider than high (28:21), subtriangular, with vertex almost straight, cheeks distinctly concave, widening towards clypeus and mandibles; interorbital space larger than eye height (45:29); frons evenly covered with delicate punctate-reticulate sculpture and scattered setae; toruli with dorsal carina forming shelf; interantennal process acute, projecting between toruli; clypeus subquadrate, medially gently concave. Antenna with scape much longer than radicle (24:7), scape micro-alutaceous. Clava 3-merous, noticeably longer than combined length of pedicle and flagellomeres 1–5.

Mesosoma distinctly longer than high (11:7), moderately convex dorsally; pronotum and mesoscutum, with delicate reticulation and scattered setae; scutellum polished, twice as wide as long; posterior margin of scutellum crenulate; dorsellum (= metascutellum) about 6 × wider than long, smooth between metascutellar carinae; sulcus separating metanotum from propodeum not raised, only a fine line; propodeum with anteriodorsal transverse concave plate from which the foamy keels arise, extending lateroventrally on each side of propodeum; lateral foamy edges are bent up like flanges; mesopleuron smooth, but with a complete longitudinal striation dorsally accompanied by a few shorter striations in posterodorsal third; transepisternal line present, anterior third arched ventrally to meet mesopleural carina; acetabular and ventral mesopleural carinae delicate; metapleuron with rather dense, long white setae except for around depression and dorsal and posterior edge forming wide foamy area. Fore wing curved over metasoma, only just surpassing tip of metasoma, 1.4× length of metasoma, very broad (15:7), without marginal cilia, with extremely short pointed white microtrichiae with bulbous base; hind wing (22:5) with distinct marginal cilia only basally.

Metasoma equal in length to rest of body, wider than high (5:2); T1 broadly trapezoidal (11:3), anteriorly covered with dense patch of long white setae; T2 distinctly wider than long (19:10), anterior margin with two broad setose depressions; T3–T6 short; S1 covered with dense patch of long, white setae.
Male. Similar to females except for fore wings of normal shape, not curved over metasoma; head with hyperoccipital carina absent between lateral ocelli and inner margin of compound eye; occiput not excavated; parapsidal lines on mesoscutum present; anterior admedian depressions more distinct than in female, pit-like; tibiae much less robust; antenna filiform.
Diagnosis
Afrisolia anyskop has a distinctly shaped head as a result of the concave genae, which make the compound eyes protrude laterally; the dorsal pronotal area is significantly expanded with well-developed pronotal shoulders, such that A. anyskop has an obvious ‘neck’, similar to Sceliotrachelus; notauli far more narrow (7× longer than wide) and parallel sided than in other species, where the notauli are at most 5× longer than wide; mesoscutal sculpturing micro-reticulate as in A. quagga, more polished in A. obesa and A. robertsoni.
Etymology
Named after the type locality, Anyskop farm. Noun in apposition.
Distribution
South Africa (Western Cape).
Afrisolia obesa
Afrisolia obesa Masner & Huggert, 1989: 35 (original description); Vlug, 1995: 10 (catalogued, type information).
Material examined
Holotype
: South Africa • ♀; Grahamstown, 24 June 1955, E. McCallan (
Diagnosis
Overall a much lighter-coloured species as in A. quagga, with a red-orange metasoma, and red-brown mesosoma and head, whereas A. anyskop and A. robertsoni have a dark brown-black body, and A. quagga has an orange-yellow metasoma contrasting strongly with the darker head and mesosoma; A. obesa has OOL shorter than LOL, whereas it is longer than LOL in A. quagga, and equal in length to LOL in A. anyskop and A. robertsoni; notauli are broader, 3× longer than wide as in A. quagga, whereas remaining species have narrower notauli, at least 5× longer than wide; mesocutum polished between notauli as in A. robertsoni, alutaceous reticulate in A. anyskop and A. quagga; only a single strong dorsal striation on mesopleuron, whereas other species have two or more striations.

Biology
Unknown.
Distribution
South Africa (Eastern Cape) (
Afrisolia quagga , sp. nov.
Material examined
Holotype
: South Africa • ♀; Eastern Cape, Blauwe Krans Farm, (12.8 km 216°SW Kirkwood) 33°30.747'S, 25°24.644'E; 9–16 Feb 2001; HG Robertson & R Tourle; Winkler; VB01-A3N-W10; Valley Bushveld (non-trashed); SAM-HYM-P095157 (

Paratypes
: South Africa • 2♂♂; E. Cape, Asante Sana Game Reserve, Waterkloof; S32 14.745 E24 56.471; 1807 m; 18–25 Nov 2009; J. Midgley; T3S3b; Pitfall trap; Camdeboo Escarpment Thicket; Mixed Grass (tussock & tall shrubs with scattered small oubos); SAM-HYM-P043318; SAM-HYM-P095174 (

Description
Female. Body length 1.1 mm. Colour of head black, mesosoma orange-brown, metasoma yellow-orange, antennae, alar sclerites, and legs yellow; wings are hyaline, but appear white due to dense presence of white microtrichiae.

Afrisolia quagga van Noort & Lahey, sp. nov. male paratype (
Head in dorsal view strongly transverse (1:3), lens-like, with frons only gently arched; temples straight, strongly receding toward occipital carina; POL:LOL:OOL = 22:10:15. Scape distinctly shorter than interorbital space (24:15) with scrobe absent; eyes glabrous; scattered white setae on head; head with delicate alutaceous reticulation, including posterior of hyperoccipital carina; head in lateral view more than twice as high as wide (9:21), with lower frons gently convex, vertex acute and posterior toruli distinctly raised; eyes oval (9:12) and upper part of gena almost disappearing behind eye; malar space much shorter than eye height (19:28); head in anterior view wider than high (12:9), lenticular, with vertex gently rounded, cheeks straight; interorbital space larger than eye height (17: 14); toruli with dorsal carina forming bilateral convex shelf, medial concave; interantennal process acute, slightly projecting between toruli. Antenna with scape to radicle as 24:5, scape micro-alutaceous. Clava 3-merous, length subequal to combined length of pedicle and flagellomeres 1–5.
Mesosoma distinctly longer than high (12:8), moderately convex dorsally; pronotum and mesoscutum, with delicate alutaceous reticulation and scattered setae; scutellum polished, more than twice as wide as long (9:4), anteromedially pointed; posterior margin of scutellum smoothly convex; dorsellum (= metascutellum) about 8× wider than long, smooth between metascutellar carinae; sulcus separating metanotum from propodeum raised; propodeum with anteriodorsal transverse concave plate from which the foamy keels arise, extending lateroventrally on each side of propodeum; lateral foamy edges are bent up like flanges; mesopleuron smooth, but with a two complete longitudinal striations dorsally accompanied by a few shorter striations in posterodorsal third; transepisternal line present, anterior third arched ventrally to meet mesopleural carina; acetabular and ventral mesopleural carinae delicate; metapleuron with rather dense, long white setae except for around depression and dorsal and posterior edge forming wide foamy area. Fore wing curved over metasoma, extending beyond tip of metasoma, 1.6× longer than length of metasoma (35:22), broad (19:10), without marginal cilia, with extremely short pointed white microtrichiae with bulbous base; hind wing (3:1) with distinct marginal cilia along full length of posterior margin.
Metasoma shorter than rest of body (11:13), wider than high (10:3); T1 broadly trapezoidal (16:5), anteriorly covered with patch of scattered long white setae; T2 distinctly wider than long (32:15), anterior margin with single transverse setose depression; T3–T6 short; S1 covered with dense patch of long, white setae.
Male. Similar to females except for fore wings of normal shape, not curved over metasoma, twice as long as metasoma; head with hyperoccipital carina less pronounced and absent between lateral ocelli and inner margin of compound eye; occiput not as laterally excavated as in females; antenna filiform; notauli much broader and more lenticular in shape, posteriorly very narrow; dorsal striations on mesopleuron more numerous. Colouration similar to female, except for pedicel and flagellum, which is dark brown.
Diagnosis
A distinctly coloured species with an orange-yellow metasoma contrasting strongly with the red-brown mesosoma and dark brown head; A. quagga has OOL longer than LOL, whereas it is shorter or equal in length to LOL in the other species; mesocutum alutaceous reticulate between notauli as in A. anyskop, polished in A. obesa and A. robertsoni; notauli are broader, 3× longer than wide as in A. obesa, whereas remaining species have narrower notauli, at least 5× longer than wide; notauli positioned closer together relative to scutellar width, meeting the mesoscutellar sulcus at least double the posterior notaular width from the lateral margins of the scutellum, in contrast to other species where the notauli are posteriorly more widely spaced.
Etymology
Named for the similar coloration and habitat affiliation to the Quagga, an extinct Plains Zebra subspecies that previously existed in the Cape region. These zebras had reduced stripes and a distinct orange hue on the flanks and hind quarters, which is also reflected in the coloration of the metasoma of this new Afrisolia species. Noun in apposition.
Distribution
South Africa (Eastern Cape).
Afrisolia robertsoni , sp. nov.
Material examined
Holotype
: South Africa • ♀; Eastern Cape, Februarie Farm, (40.2 km 267°W Kirkwood); 33°33.124'S, 25°03.043'E; 10–12 Feb 2001; S. van Noort; Malaise trap; VB01-R1N-M22; Valley Bushveld (non-trashed); SAM-HYM-P095158 [OSUC 20243869] (
Description
Female. Body length 0.95 mm. Colour of body black, with clava, metasoma and alar sclerites dark brown; rest of antennae, all legs and mandibles yellow-brown; wings basally appear white due to dense presence of white microtrichiae, appear infuscate over distal two-thirds due to dark microtrichae.

Afrisolia robertsoni van Noort & Lahey, sp. nov. female holotype (
Head in dorsal view strongly transverse (5:15), lens-like, with frons only gently arched; temples straight, strongly receding toward occipital carina; POL:LOL:OOL = 8:5:5. Scape distinctly shorter than interorbital space (38:57) with scrobe absent; eyes with minute hairs, scattered setae on head slightly longer; head with delicate alutaceous reticulation, including posterior of hyperoccipital carina; head in lateral view about twice as high as wide (12:25), with lower frons gently convex, vertex acute and posterior toruli distinctly raised; eyes oval (5:8) and upper part of gena almost disappearing behind eye; posterior edge of gena with long, posteriorly projecting setae from ventral section; malar space much shorter than eye height (40:75); head in anterior view wider than high (20:13), lenticular, with vertex gently rounded, cheeks straight; interorbital space larger than eye height (6: 4); frons evenly covered with delicate punctate-reticulate sculpture and scattered setae; toruli with dorsal carina forming medially convex shelf; interantennal process acute, projecting between toruli. Antenna with scape to radicle as 75:20, scape micro-alutaceous. Clava 3-merous, noticeably longer than combined length of pedicle and flagellomeres 1–5.
Mesosoma distinctly longer than high (17:12), moderately convex dorsally; pronotum and mesoscutum with delicate reticulation and scattered setae; scutellum polished, more than twice as wide as long (9:4), anterior margin medially pointed; posterior margin of scutellum smoothly convex; dorsellum (= metascutellum) about 7 × wider than long, smooth between metascutellar carinae; sulcus separating metanotum from propodeum not raised, only as fine line; propodeum with anteriodorsal transverse concave plate from which the foamy keels arise, extending lateroventrally on each side of propodeum; lateral foamy edges are bent up like flanges; mesopleuron smooth, but with a complete longitudinal striation dorsally accompanied by a few shorter striations in posterodorsal third; transepisternal line present, anterior third arched ventrally to meet mesopleural carina; acetabular and ventral mesopleural carinae delicate; metapleuron with rather dense, long white setae except for around depression, dorsal and posterior edge forming wide foamy area. Fore wing straight, not curved over metasoma, extending well beyond tip of metasoma, 1.7× longer than length of metasoma (19:11), broad (19:10), without marginal cilia, with extremely short pointed white microtrichiae with bulbous base; hind wing (30:7) with distinct marginal cilia only basally.
Metasoma equal in length to rest of body, wider than high (4:2); T1 broadly trapezoidal (12:3), anteriorly covered with dense patch of long white setae; T2 distinctly wider than long (17:10), anterior margin with two broad setose depressions; T3–T6 short; S1 covered with dense patch of long, white setae.
Male. Unknown.
Diagnosis
Afrisolia robertsoni is overall a dark species as in A. anyskop, opposed to the much lighter A. obesa and A. quagga, which have distinctly paler metasomas; OOL is equal in length to LOL as in A. anyskop, shorter than LOL in A. obesa, longer than LOL in A. quagga; the straight genae separate this species from A. anyskop, which has a distinct head-shape as a result of the concave genae; notauli broader (5× longer than wide) than in A. anyskop (7× longer than wide), and narrower than in A. obesa and A. quagga (3× longer than wide); mesocutum polished between notauli as in A. obesa, alutaceous reticulate in A. anyskop and A. quagga.
Etymology
Named after Hamish Robertson, previous director and entomologist at the Iziko South African Museum, colleague and friend of Simon van Noort. Together they implemented the entomological sphere of the Conservation Farming Project through which the type specimen was procured. Noun in the genitive case.
Distribution
South Africa (Eastern Cape).
Allotropa
Allotropa
Förster, 1856: 106, 109 (original description. Type: Inostemma mecrida Walker, by monotypy. Keyed);
Eurostemma
Szelényi, 1938: 102 (original description. Type: Inostemma europus Walker, by monotypy and original designation. Synonymized by Vlug and Graham (1984));
Nasdia
Nixon, 1942: 462 (original description. Type: Nasdia prosper Nixon, by monotypy and original designation. Synonymized by
Platytropa
Kozlov, 1977: 79, 80 (original description. Type: Platytropa helenae Kozlov, by monotypy and original designation. Keyed. Synonymized by
Diagnosis
Head in dorsal view wider than long, subellipsoidal to lens-like. Occipital pit not developed. Temples very short to almost absent. Posterior ocellus at most 1 diameter distant from inner orbit; OOL<LOL. Facial and malar striae absent. Mandibles long, bidentate and crossing scissor-like. Female antenna with semi-abrupt 3-merous clava; male antenna often with whorls of long, erect bristles on A3–A7. Mesosoma with notauli absent. Scutoscutellar sulcus foveolate. Propodeum centrally with elevated, glabrous central keel or triangular bulge. Netrion not developed. Transepisternal line deep and horseshoe-shaped; metapleuron and sides of propodeum hairy. Tarsal formula 5-5-5 (4-4-4 in one undescribed species). T1 transverse, longitudinally costate. Metasoma with lateral edge indistinct.


Species richness
Allotropa canopyana Buhl, 2011 (Tanzania).
Allotropa delottococci Buhl, 2019 (South Africa. Introduced to Spain).
Allotropa fusca Buhl, 2011 (Tanzania).
Allotropa kamburovi Annecke & Prinsloo, 1977 (South Africa) (Fig.
Allotropa loundsburyi Ashmead, 1901 (South Africa).
Allotropa magnini Risbec, 1955 (Ivory Coast).
Allotropa pauliana Risbec, 1955 (Madagascar).
Allotropa species (South Africa) (Fig.
Distribution
Afrotropical: Ivory Coast, Madagascar, South Africa, Tanzania. Cosmopolitan, excluding Antarctica and New Zealand (
Biology
Solitary and gregarious endoparasitoids of mealybugs and cochineals (Hemiptera, Sternorrhyncha, Pseudococcidae and Dactylopiidae).
Allotropa canopyana
Allotropa canopyana Buhl, 2011: 76 (original description, illustrated).
Material examined
Holotype
: Tanzania • ♀; Tanga, Lushoto Dist., Mazumbai For. Res.; 1370–1435 m; 4.xii.1995 (
Biology
Unknown.
Distribution
Tanzania (
Allotropa delottococci
Allotropa delottococci Buhl, 2019: 60 (original description, species key, illustrated).
Material examined
Holotype
: South Africa • ♀; Limpopo province, Letsitele; 7358977'S, 239175'E UTM coordinates; grid 36K; 14.vi.2017; Marco Benito; from Delottococcus aberiae (De Lotto) in crop of orange Citrus sinensis (L). Osbeck (
Paratypes
: South Africa • 3♀♀, 2♂♂ same data as holotype (
Biology
Parasitoid of the mealybug Delottococcus aberiae (De Lotto, 1961) (Hemiptera, Pseudococcidae) (
Distribution
South Africa. Introduced into Spain as a biocontrol agent of Delottococcus aberiae (
Allotropa fusca
Allotropa fusca Buhl, 2011: 77 (original description, illustrated).
Material examined
Holotype
: Tanzania • ♀; Tanga, Muheza Dist., Kwamgumi For. Res.; 170–220 m; 25.vii.1995 (
Biology
Unknown.
Distribution
Tanzania (
Allotropa kamburovi
Allotropa kamburovi
Annecke & Prinsloo, 1977: 105 (original description, illustrated);
Material examined
Holotype : South Africa • ♀; Pretoria, Tvl., laboratory culture. S. Kamburov, ex Planococcus citri (Risso) on potato sprouts. Type material was held in culture for a number of months having first emerged from P. citri collected at Rustenburg, Tvl., ii. 1976, by E.C.G. Bedford (T 5025) (SANC).
Paratypes
: South Africa • 1♂; same data as holotype; slide mounted (SANC); 24♀♀, 43♂♂, same data as holotype (
Comments
Only a single male is present in the
Biology
Solitary endoparasitoid of the citrus mealybug Planococcus citri (Risso) on potato sprouts (
Distribution
South Africa (
Allotropa loundsburyi
Allotropa loundsburyi
Ashmead, 1901: 138 (original description);
Material examined
Holotype
: South Africa • ♀; Cape Colony; 22 Oct. 1898; C.P. Lounsbury; ex Dactylopius sp. on gorse; Cat. No. 5727 (
Comments
The
Biology
Parasitoid of Dactylopius sp. (Hemiptera, Dactylopiidae) on gorse (
Distribution
South Africa (
Allotropa magnini
Allotropa magnini
Risbec, 1955a: 234 (original description, illustrated);
Material examined
Syntypes : Ivory Coast • 39♀♀, 50♂♂; Bingerville; vi–vii.1953; Magnin; Allotropa magnini Risbec; ex larves de Pseudococcus njalensis (MNHN).
Biology
Parasitoid of Formicococcus njalensis (Laing, 1929) (Hemiptera, Pseudococcidae) (
Distribution
Ivory Coast (
Allotropa pauliana
Allotropa pauliana
Risbec, 1955b: 117 (original description, biology, illustrated);
Allotropa pauliani
Risbec:
Material examined
Syntypes : Madagascar • 2♀♀, 1♂, 2 unsexed adults within host remains (MNHN).
Biology
Parasitoid of mealybugs on Philippia species (Risbec 1955).
Distribution
Madagascar (
Amitus
Amitus
Haldeman, 1850: 109 (original description. Type: Amitus aleurodinis Haldeman, by monotypy);
Zacrita
Förster, 1878: 46 (original description. Type: Zacrita longicornis Förster, by monotypy and original designation. Synonymized by
Elaptus
Forbes, 1885: 110 (original description. Type: Elaptus aleurodis Forbes, by monotypy. Error for Alaptus Westwood. Synonymized implicitly by
Passalida
Brèthes, 1914: 2 (original description. Type: Passalida spinifera Brèthes, by monotypy and original designation. Synonymized by
Masnerium
Polaszek, 2009: 120 (original description. Type: Masnerium wellsae Polaszek, by monotypy and original designation). Synonymized by
Diagnosis
Short, stocky, dorsoventrally flattened species, with long wings without distinct veins; head in lateral view somewhat opisthognathous; antennal formula usually 10-10, rarely 8-8 (e.g. Amitus wellsae (Polaszek)); female antenna with abrupt, subcompact, spindle-shaped to ovoid clava resulting from the fusion of A8–A10; sulci present between clavomeres except in Australian members of the genus (e.g. Amitus wellsae (Polaszek)); male antenna with specialized paddle-shaped area on A4; netrion well developed; propodeum partly covered with foamy structures; metasoma short, subsessile, almost as wide as long; T1 strongly trapezoidal-transverse; T2 usually with fan of striae anterolaterally.


Species richness
Amitus hesperidum Silvestri, 1927 (China, but introduced to most tropical areas) (Fig.
Amitus species 1 (Kenya).
Amitus species 2 (Madagascar).
Amitus species 3 (South Africa) (Fig.
Distribution
Afrotropical: Kenya, Madagascar, South Africa (new records). Cosmopolitan, excluding Antarctica and New Zealand (
Biology
Solitary and occasionally gregarious endoparasitoids of whiteflies (Hemiptera, Sternorrhyncha, Aleyrodidae) (
Amitus hesperidum
Amitus hesperidum
Silvestri, 1927: 55 (original description);
Amitus hesperidum variipes Silvestri, 1927: 58 (original description).
Amitus variipes
Silvestri, 1927:
Material examined
Holotype
: China • Lost. Redescribed by
Additional material examined
USA • 2♀♀, 1♂ card mounted; 13♀♀, 2♂♂ loose in gelatine capsule; Florida, Gainsville; xi. 1989; Ru Nguyen; ex Aleurocanthus woglumi on citrus; ex consignment; AcBC 275; Amitus hesperidum (SANC).
Biology
Parasitoid of the citrus blackfly, Aleurocanthus woglumi (Hemiptera, Aleyrodidae), a major pest of citrus trees (
Distribution
China, but introduced to most tropical areas of the world as part of biocontrol programs (
Comments
No published evidence of introduction or establishment of A. hesperidum in South Africa could be located, possibly because Eretmocrus serius Silvestri (Chalcidoidea, Aphelinidae) was introduced and successfully controlled citrus blackfly after its discovery in 1959 in South Africa (
Fidiobia
Fidiobia
Ashmead, 1894: 170 (original description. Type: Fidiobia flavipes Ashmead, by monotypy);
Rosneta
Brues, 1909: 157 (original description. Type: Rosneta tritici Brues, by monotypy and original designation. Synonymized by
Fidobia
Ashmead, 1894:
Triclavus
Brèthes, 1916: 411 (original description. Type: Triclavus bonaeriensis Brèthes, by monotypy. Synonymized by Masner, in
Fahringeria
Kieffer, 1921: 68 (original description. Type: Fahringeria synergorum Kieffer, by monotypy. Synonymized by
Platyllotropa
Szelényi, 1938: 126 (original description. Type: Platyllotropa gallicola Szelényi, by monotypy and original designation. Synonymized with Triclavus Brèthes by Ogloblin (1944));
Diagnosis
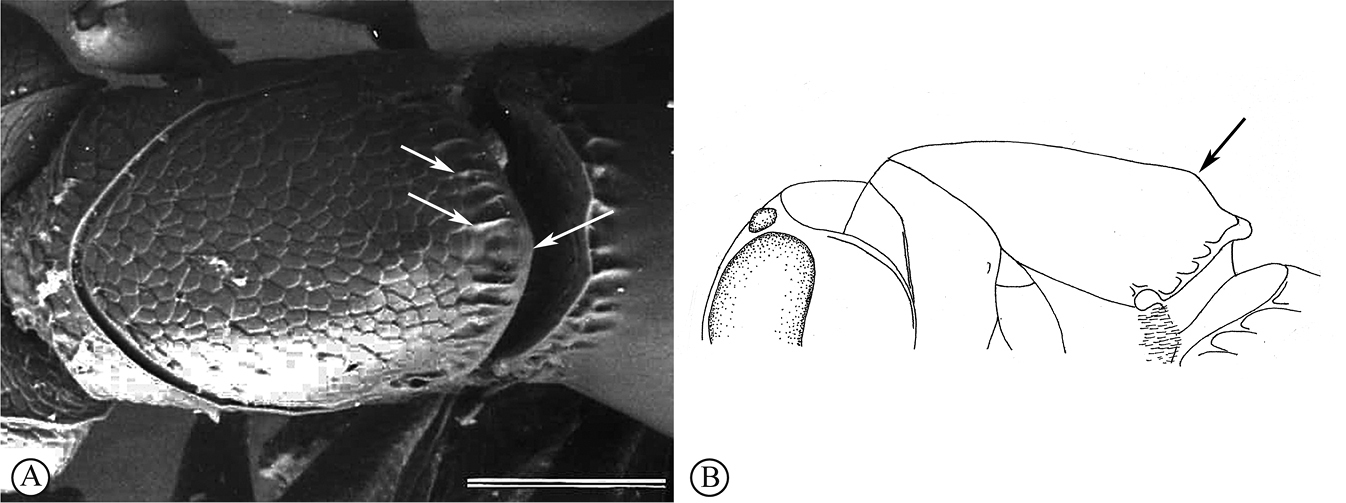
Minute species (0.6–1.3 mm) with body slightly to considerably depressed dorsoventrally; mostly melanic, with brightly coloured appendages; vertex rounded without hyperoccipital carina. OOL variable, but in most species very short, equal to or shorter than diameter of posterior ocellus; antenna 9- or 10-merous, in females with abrupt, 3-merous clava; A8–A10 slightly less abrupt in males. Mesoscutum flattened; notauli (if present) abbreviated anteriorly, gradually dilated posteriorly. Fore wing in most species with short tubular submarginal vein. T2 the largest tergite, with two depressions anterolaterally (
Species richness
Fidiobia benjamini (Nixon, 1969) (Kenya) (Fig.
Fidiobia celeritas van Noort & Lahey, sp. nov. (South Africa) (Figs
Fidiobia danielssoni Buhl, 2001 (South Africa) (Figs
Fidiobia filicornis Buhl, 2014 (Togo).
Fidiobia semirufa Buhl, 2014 (Togo).
Fidiobia tanzaniana Buhl, 2010 (Tanzania).
Fidiobia tschirnhausi Buhl, 2014 (Togo).
Fidiobia zebra Buhl, 2010 (Tanzania).
Fidiobia species (Tanzania) (Figs
Distribution
Afrotropical: Kenya, South Africa, Tanzania, Togo. Cosmopolitan, excluding Antarctica (
Biology
Solitary endoparasitoids of weevil (Coleoptera, Curculionidae) and leaf beetle (Coleoptera, Chrysomelidae) eggs (
Comments
There are numerous further undescribed species of Fidiobia from the Afrotropical region present in the collections of
Fidiobia benjamini
Platystasius benjamini Nixon, 1969: 445, 449 (original description, keyed).
Fidiobia benjamini
(Nixon):
Material examined
Holotype
: Kenya • ♀; Kangaita; 26.i.1967; ex Entypotrachelus micans on tea; CIEA2853 (

Paratypes
: Kenya • 2♀♀; same data as holotype (
Biology
Egg parasitoid of Entypotrachelus micans Hustache, 1929 (Coleoptera, Curculionidae) on tea, Camellia sinensis (
Distribution
Kenya (
Fidiobia celeritas , sp. nov.
Material examined
Holotype
: South Africa • ♀; Eastern Cape, Februarie Farm, (40.2 km 267°W Kirkwood) 33°33.124'S, 25°03.043'E; 10–12 Feb 2001; S. van Noort; Malaise trap; VB01-R1T-M21; Valley Bushveld (goat trashed); SAM-HYM-P095159 (
Paratypes
: South Africa • 1 ♂; same data as holotype except for SAM-HYM-P095160 (
Description
Female. Body length 0.9 mm. Colour of body dark brown to black, with metasoma, antennal funicular segments, alar sclerites brown; scape and pedicel and legs yellow; wings clear with brown microtrichiae.

Head in dorsal view strongly transverse (5:13), lens-like, with frons only gently arched; temples straight, strongly receding toward occipital carina; POL:LOL:OOL = 40:22:20. Scape distinctly shorter than interorbital space (7:10) with scrobe absent; eyes with minute hairs, scattered white setae on head slightly longer; head with delicate alutaceous reticulation; head in lateral view two-thirds as wide as high (10:15), with lower frons gently convex, vertex acute and posterior toruli distinctly raised; eyes oval (7:10) with upper part of gena still visible behind eye; posterior edge of gena defined by crenulate and sharp occipital carina with c. 5 long, posteriorly projecting setae from ventral section of carina; malar space over half as long as eye height (55:83); head in anterior view wider than high (14:10), broadly oval, with vertex rising to ocelli, cheeks gently convex; interorbital space larger than eye height (80:63); frons evenly covered with delicate punctate-reticulate sculpture and scattered setae; toruli with dorsal carina not forming shelf; interantennal process absent; clypeus ellipsoidal, gently convex in profile, anterior margin distinctly convex. Antenna with 10-antennomeres, with scape to radicle as 11:2, scape micro-alutaceous. Clava 3-merous, equivalent in length to combined length of pedicle and flagellomeres 1–5.

Mesosoma distinctly longer than high (14:10), wider than long (95:80); moderately convex dorsally; pronotum and mesoscutum, with delicate reticulation and scattered white setae; scutellum polished, twice as wide as long; posterior margin of scutellum crenulate; dorsellum (=metascutellum) medially hidden in dorsal view; sulcus separating metanotum from propodeum not raised, only as fine line; propodeum with an anteriodorsal transverse flat, polished plate from which the parallel foamy keels arise, extending lateroventrally on each side of propodeum forming an H-shape; lateral foamy edges extend as moderate flanges; mesopleuron smooth, but dorsally with a set of 14–15 longitudinal striations, the dorsal five transversely complete, reaching the anterior mesopleural edge, subsequent ventral striations shorten sequentially towards the transepisternal line; transepisternal line present, anterior third arched ventrally to meet mesopleural carina; acetabular and ventral mesopleural carinae delicate; metapleuron with dense, long white setae except for around depression and dorsal and posterior edge forming wide foamy keels. Fore wing only slightly curved over metasoma, well surpassing tip of metasoma, 1.7× length of metasoma, very broad (2:1), with very short marginal cilia, and extremely short pointed brown microtrichiae with slightly bulbous base; submarginal vein ending with distinct rounded club, just over a quarter (0.27×) of fore wing length; hind wing (5:1) with distinct marginal cilia apically and basally.

Metasoma equal in length to rest of body, wider than high (15:6.5); T1 broadly trapezoidal (4:1), anterolaterally covered with sparse patches of long white setae; anteromedially with two depressions filled with short white setae, one each side of the nucha which has a raised carina; T2 distinctly wider than long (15:8), anterior margin with two transversely narrow setose depressions; T3–T6 short; ovipositor apically serrate.
Male. Similar to females except for fore wings of normal flat shape, not curved over metasoma, narrower, twice as long as wide; legs brown; antenna filiform, almost as long as body, with 9 antennomeres, flagellar segments (A3–A9) of equivalent length (0.5× scape length), very setose with short, freely projecting multiporous plate sensillae present in 4–5 staggered rows over entire funicle segment.
Etymology
Named after the Latin word for speed with reference to the notauli and median sulcus configuration on the mesoscutum that is reminiscent of GT racing stripes. Noun in apposition.
Diagnosis
Both sexes are immediately distinguishable by the presence of a median mesoscutal line, which is absent in the other species. The notauli extend far forward, almost meeting the admedian depressions. Males are unique amongst other members of the genus, and the subfamily Sceliotrachelinae, by possessing the sex segment (tyloid) on A3 because of the fusion of A3 and A4, resulting in 9-merous antennae in the male.
Biology
Unknown.
Distribution
South Africa (Eastern Cape).
Comments
This species strongly resembles the genus Afrisolia, so much so that it was misidentified as a member of that genus in figure 3 of
Fidiobia danielssoni
Fidiobia danielssoni Buhl, 2001: 22 (original description).
Material examined
Holotype : South Africa • ♀; Cape Province, Koomplanskloof, 10 km S Citrusdal; 200–270 m; 32°40'S, 19°01'E; 4–8.X.1994; Malaise trap; R. Danielsson (MZLU).

Fidiobia danielssoni Buhl, 2001 holotype (MZLU) (A, B) and Fidiobia species (Tanzania) (
Biology
Unknown.
Distribution
South Africa (Buhl 2001).
Fidiobia filicornis
Fidiobia filicornis Buhl, 2014: 74, 75, 76 (original description, illustrated).
Material examined
Holotype
: Togo • ♂; Région des Plateaux, Cascade d’Ayomé NE of Amlamé; 07°30'08"N, 00°57'20"E; 305–330 m; 13.iv.2008; at shady creek bank in rock gorge; much Anubias gigantea; swept; M. von Tschirnhaus (

Biology
Unknown.
Distribution
Togo (Buhl 2014).
Fidiobia semirufa
Fidiobia semirufa Buhl, 2014: 75, 76 (original description, illustrated).
Material examined
Holotype
: Togo • ♂; Région des Plateaux, Zogbégan, village part Zogbégan-Carriére (SE of Badou), at creek Elèbè, V-shaped valley near cocoa plantation downstream of village; 07°34'50"N, 00°40'03"E; 20–25.iv.2008; 650 m; remains of secondary rainforest; swept; M. von Tschirnhaus (
Biology
Unknown.
Distribution
Togo (Buhl 2014).
Fidiobia tanzaniana
Fidiobia tanzaniana Buhl, 2010: 33 (original description, illustrated).
Material examined
Holotype
: Tanzania • ♀; Udzungwa Mts, Iringa Region, Kilolo dist., Ndundulu Forest, Matumbo camp area; 1430 m; 10–24.vii.2007; Malaise trap in semi-evergreen virgin forest; L.A. Hansen (
Biology
Unknown.
Distribution
Tanzania (Buhl 2010).
Fidiobia tschirnhausi
Fidiobia tschirnhausi Buhl, 2014: 76, 77, 78 (original description, illustrated).
Material examined
Holotype
: Togo • ♀; Région des Plateaux, Ouvêtsévé near Kpélé Élé; 07°21'27"N, 00°51'12"E; 345 m; 15.iv.2008; creek bank within forest, diverse herb vegetation; swept; M. von Tschirnhaus (
Biology
Unknown.
Distribution
Togo (Buhl 2014).
Fidiobia zebra
Fidiobia zebra Buhl, 2010: 34 (original description, illustrated).
Material examined
Holotype
: Tanzania • ♀; Udzungwa Mts., Iringa Region, Kilolo dist., Ndun-dulu Forest, Luwala camp area; 1880 m; 1–14.ii.2007; Malaise trap in semi-evergreen tropical montane virgin forest; L.A. Hansen (
Biology
Unknown.
Distribution
Tanzania (Buhl 2010).
Isolia
Isolia
Förster, 1878: 46 (original description. Type: Isolia foersteri Szabó, designated by
Diagnosis
Robust, usually dark-coloured species. OOL subequal to LOL. Frons above toruli without transverse ledge; interantennal process moderately developed. Mandibles short, strong with lower edge upcurved apically. Female antenna with abrupt 3-merous clava. Male antenna filiform. Pronotal shoulders well-developed. Notauli abbreviate anteriorly. Scutellum broadly transverse, subrectangular, with scutellaxillar pits reduced to points, and scutellar rim not defined. Mesopleuron without transepisternal line. Propodeum with foamy structures. Fore wing with no tubular veins, and with microtrichia in the form of minute, semi-erect spiculae. Fore tibial spur combed. Metasoma short and broad, with laterotergites wide and no felt fields on S2 (

Isolia hispanica female (OSUC) (A–D OSUC 20666354 E, F OSUC 20666355) A habitus, dorsolateral view B habitus, dorsal view C habitus, ventrolateral view D head, anterior view E habitus, lateral view F habitus, ventral view.
Species richness
Isolia hispanica Buhl, 1999 (Kenya). Also present in the Palearctic (Spain) (Fig.
Isolia species (Madagascar).
Biology
Unknown.
Distribution
Afrotropical: Kenya, Madagascar. Palearctic: France, Greece, Iran, Israel, Italy, Mongolia, Montenegro, Spain, Thailand, Turkey. Indomalayan (Oriental): China, India, Philippines (
Isolia hispanica
Isolia hispanica Buhl, 1999: 13 (original description).
Material examined
Holotype : Spain • ♀; Zaragoza, Los Monegros region; UTM 30TYL2794; 25.vii.1992; leg. Javier Blasco-Zumeta; Wilkening trap placed in the branches of Juniperus thurifera L.
Biology
Unknown.
Distribution
Kenya (new country record here). Also present in the Palearctic (Spain) (
Parabaeus
Parabaeus
Kieffer, 1910: 294 (original description). Type: Parabaeus ruficornis Kieffer, by monotypy and original designation);
Diagnosis
Body shape variable, from stocky and highly convex to elongate, spindle-like. All Old World species are apterous, as are the described Neotropical species with some undescribed New World species being micropterous or full-winged. Mostly yellow or light brown. Posterior ocellus contiguous with inner orbit; ocellar triangle high. Cheek and postgena with deep longitudinal excavation for housing of scape. Antennal clava of both sexes ovoid, 4-merous. Mesosoma of flightless species subrectangular, with most sclerites fused. Fore wing (when present) with short rudiment of submarginal vein without apical knob. Metasoma highly convex both dorsally and ventrally. T1 fused with T2, and S1 with S2, into solid sclerite; felt fields absent from S2 (
Species richness in the Old World
Parabaeus abyssus Austin, 1990 (Australia) (Fig.
Parabaeus africanus Austin, 1990 (Malawi)
Parabaeus armadillus Austin, 1990 (South Africa) (Figs
Parabaeus austini Buhl, 2011 (Tanzania)
Parabaeus brevicornis Buhl, 2011 (Tanzania)
Parabaeus nasutus van Noort, sp. nov. (South Africa) (Figs
Parabaeus papei Buhl, 2011 (Tanzania)
Parabaeus peckorum Austin, 1990 (South Africa)
Parabaeus quasimodus Austin, 1990 (Kenya)
Parabaeus ruficornis Kieffer, 1910 (Seychelles) (Fig.
Distribution
Afrotropical: Kenya, Madagascar, Malawi, Seychelles, South Africa, Tanzania (
Biology
Unknown. Predicted to be living near the ground, possibly as leaf-litter inhabitants (
Comments
The Old World species are all apterous, as are the described Neotropical species: P. lenkoi de Santis, 1970 (Brazil) and P. kiefferi de Santis, 1970 (Argentina), but a number of New World species are known that are also micropterous or fully winged (
Sexual dimorphism is slight in some species with morphological differences only apparent in the shape of the antennal club (
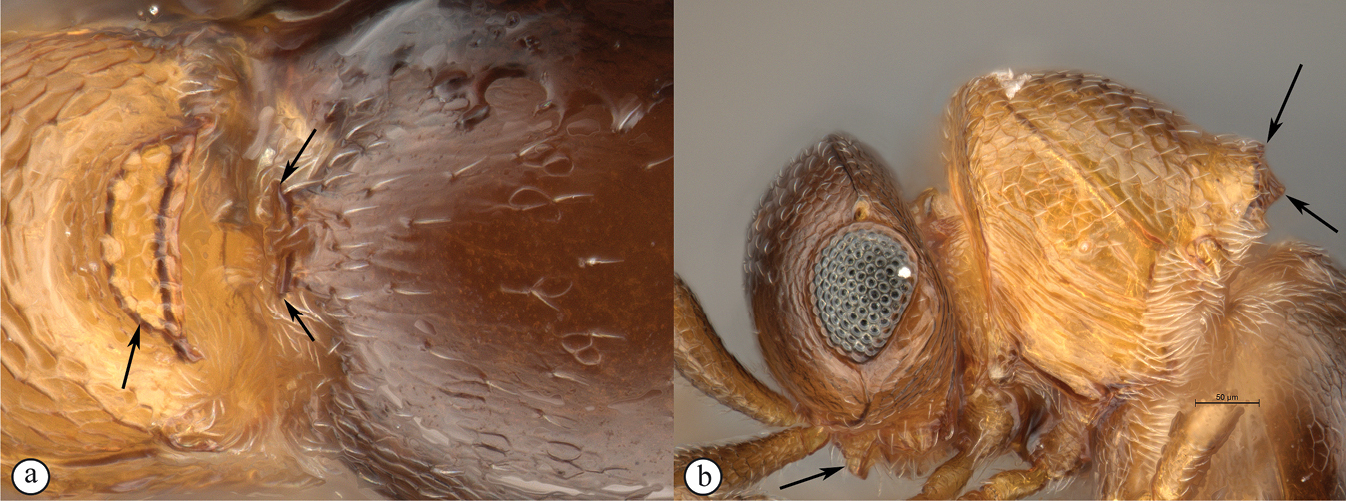
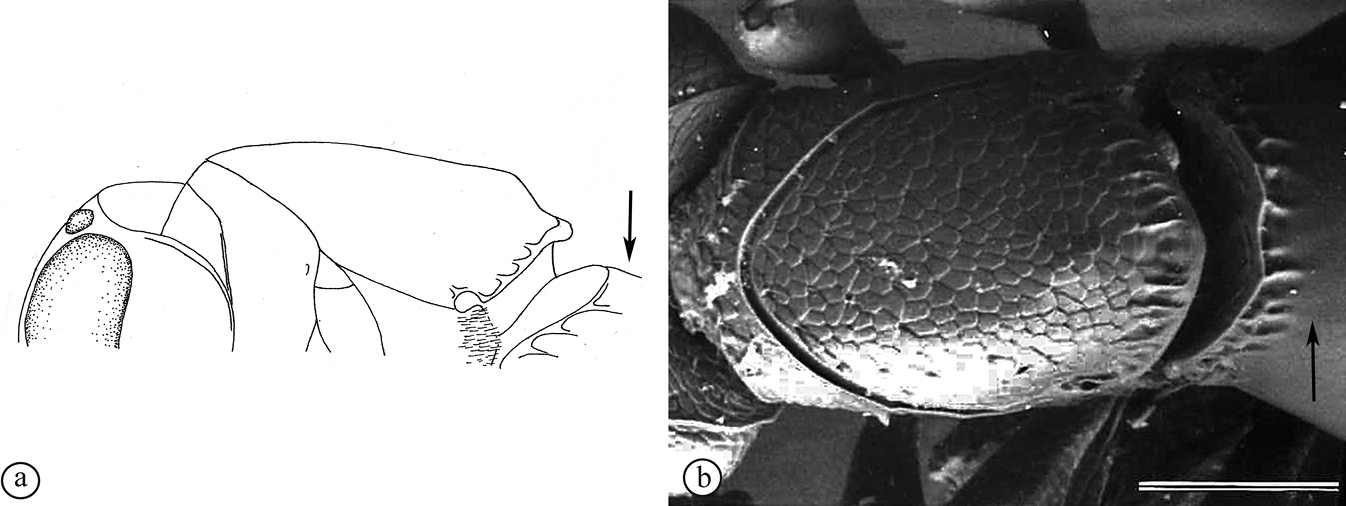
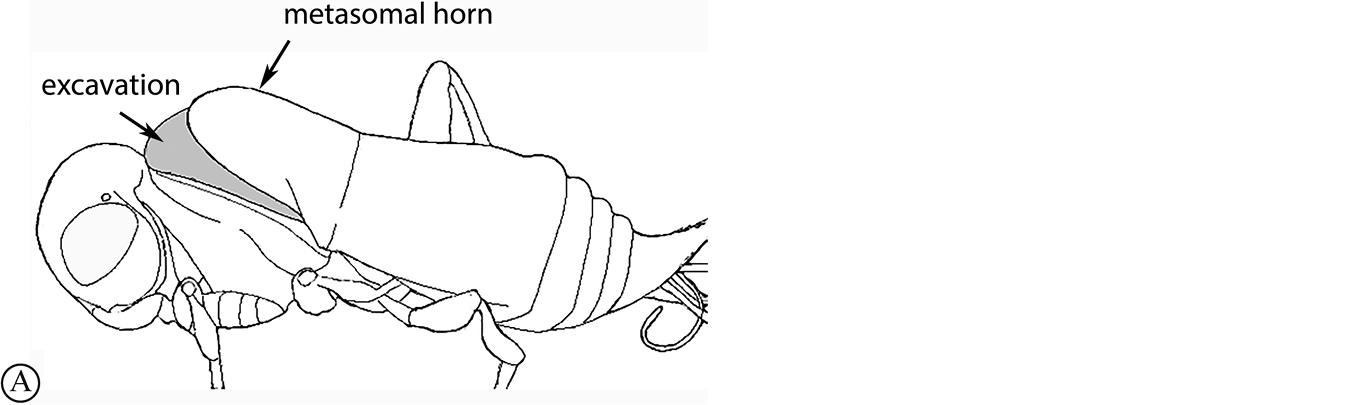
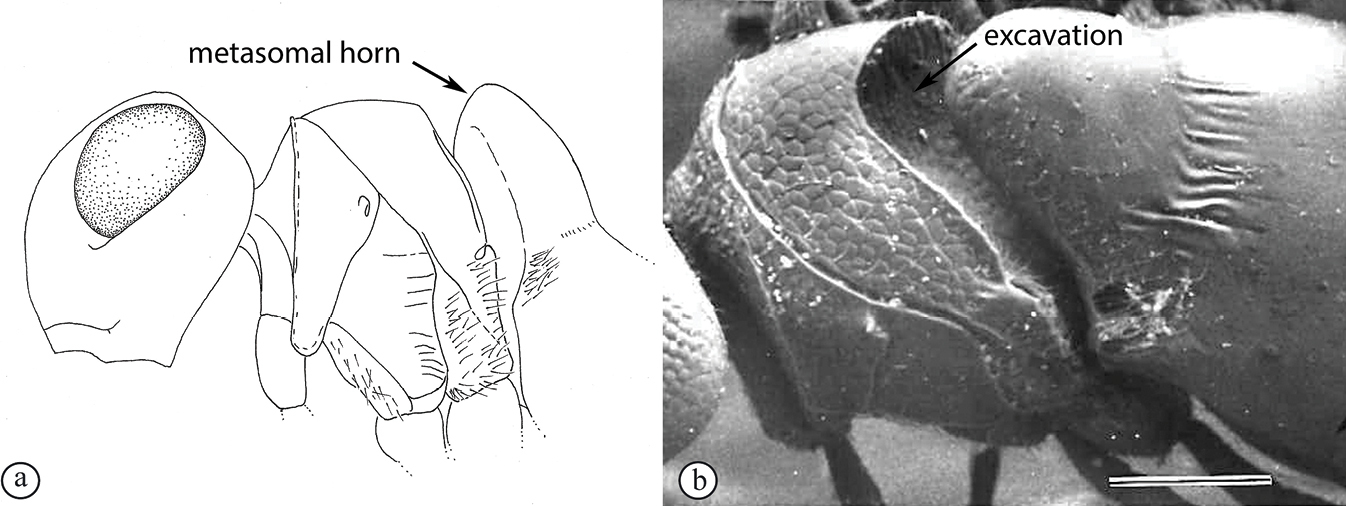
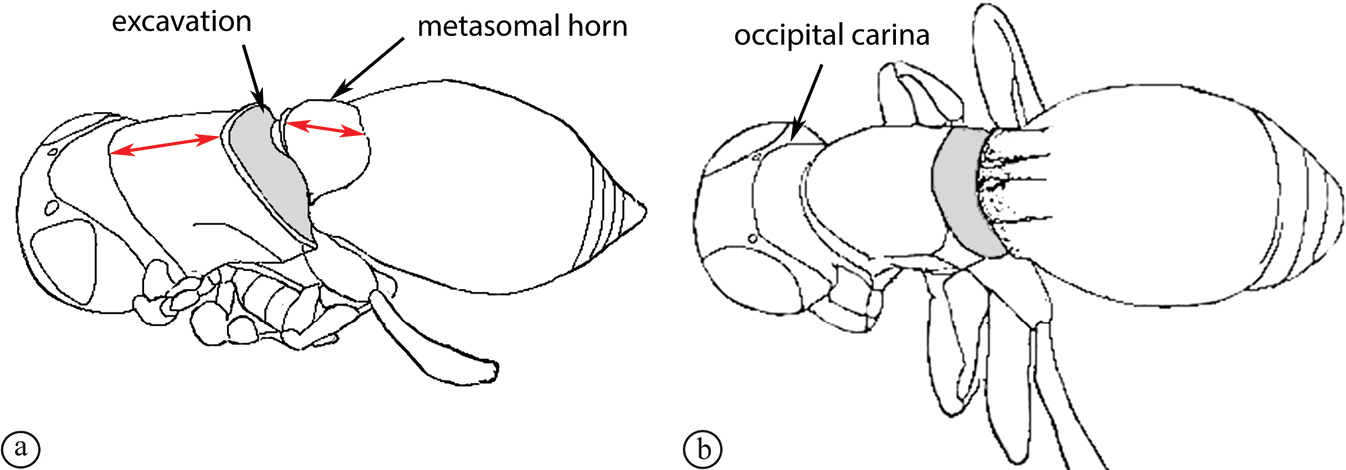
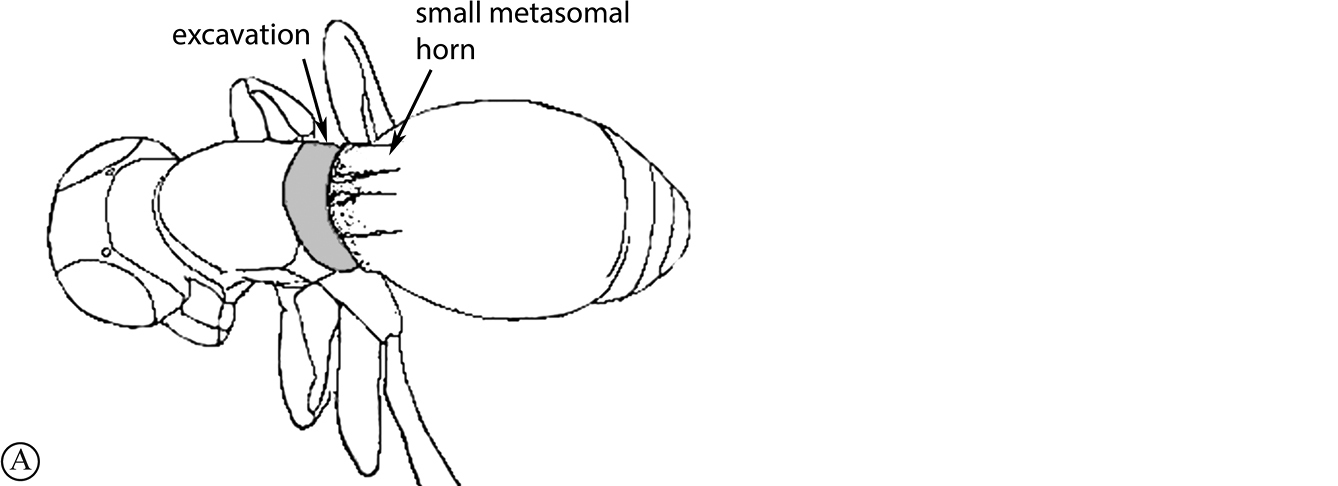
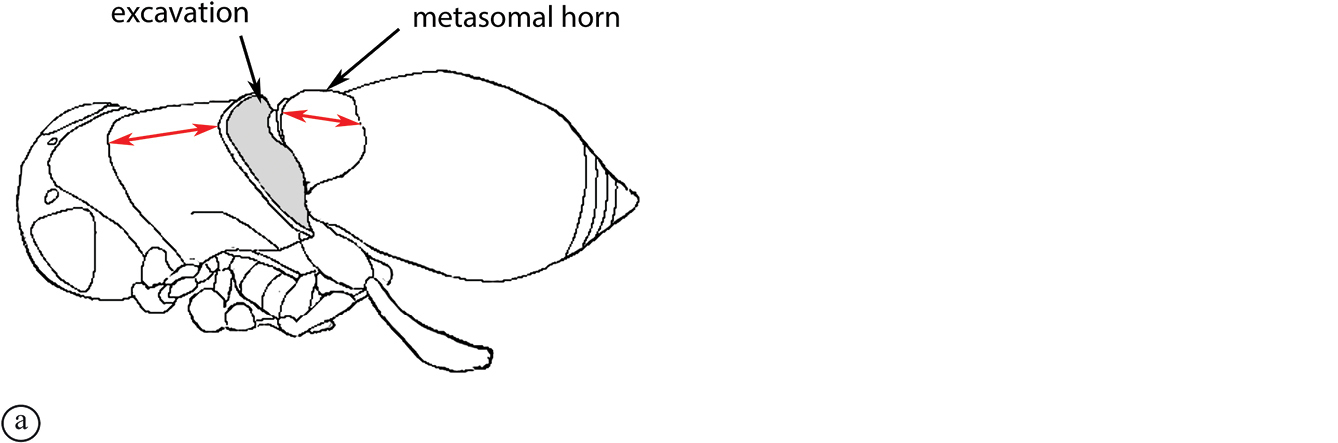
There are two apparent species-groups in the Afrotropical region defined by the presence or absence of a hyperoccipital carina. We predict that these two groups will be further supported by the presence or absence of a metasomal horn in females, once both sexes of the known species are discovered.
Parabaeus armadillus species-group (P. armadillus, P. nasutus, P. peckorum)
- Hyperoccipital carina present.
- Sexual dimorphism slight, females without metasomal horn on T1.
- Absence of a sulcus between the lateral pronotum and mesopleuron.
Parabaeus quasimodus species-group (P. africanus, P. austini, P. brevicornis, P. papei, P. quasimodus, P. ruficornis)
- Hyperoccipital carina absent.
- Sexual dimorphism strong, females with metasomal horn on T1 that is developed to varying degrees in size.
- Sulcus between the lateral pronotum and mesopleuron present.
The only other described Old World species, the Australian P. abyssus falls into its own species-group, sharing characters across the two Afrotropical species-groups (hyperoccipital carina absent, but no metasomal horn on T1 in females) and the Neotropical species-group, which has armature (points, spikes or truncate projections) on the posterior or posterolateral margin of the propodeum, and these are also present in P. abyssus (
The following key includes diagnostic characters enabling both sexes to be keyed out where known. Males of four species (P. austini, P. brevicornis, P. quasimodes, P. papei) with metasomal horns in females are as yet unknown, and hence will not be identifiable using the current key configuration.
Key to Old World species of Parabaeus
(modified after
| 1 | Hyperoccipital carina present between the lateral ocelli (distinct from occipital carina) (A, B) | 2 |
|
||
| – | Hyperoccipital carina absent, if carina present between ocelli then clearly part of occipital carina (a, b) | 4 |
|
||
| 2 | Head and mesosoma covered with small compact plates (A, B, C) | Parabaeus armadillus Austin |

|
||
| – | Head and mesosoma covered with imbricate (squamate) sculpturing (a) | 3 |
|
||
| 3 | Base of metasoma with obvious foveate pits (A); occipital carina strong and visible in dorsal view (A); mesoscutum and scutellum evenly convex (B) | Parabaeus peckorum Austin |
|
||
| – | Base of metasoma without obvious foveate pits, with bifurcate projecting medial plate (a); occipital carina not visible dorsally; posterior mesoscutum and scutellum raised into a medial plateau-like (a, b), transversely ellipsoidal projection (a); clypeus produced into nasute-like process (b) | Parabaeus nasutus van Noort, sp. nov. |
|
||
| 4 | Anterolateral mesosoma with large pit (A); dorsal mesosoma longitudinally striate (B); metasoma normal, tergite 1 not developed into a horn (B) | Parabaeus abyssus Austin |

|
||
| – | Anterolateral mesosoma without large pit (a); dorsal mesosomal surface reticulate-coriaceous (may be entirely excavated) (b); tergite 1 in females developed into a horn | 5 |

|
||
| 5 | Metasomal horn present (females) | 6 |

|
||
| – | Metasomal horn absent (males) | 9 |
|
||
| 6 | Excavation encompasses entire dorsal length of mesoscutum (A); metasomal horn extending anteriorly over three-quarters of mesosoma (A) | Parabaeus austini Buhl |
|
||
| – | Mesoscutum with dorsal surface present, excavation restricted to posterior half of mesoscutum (a, b); metasomal horn at most extending over posterior half of mesosoma (a, b) | 7 |
|
||
| 7 | Metasomal horn longer than dorsal surface of mesosoma (A, B); occipital carina low on posterior head; vertex smoothly rounded (B) | Parabaeus quasimodus Austin |

|
||
| – | Metasomal horn shorter than dorsal surface of mesosoma (a); occipital carina high on posterior head (a, b); vertex interrupted by occipital carina (a, b) | 8 |
|
||
| 8 | Metasomal horn barely developed, represented by raised area with longitudinal carinae (A, B), shallow excavation restricted to posterior face of mesosoma (A, B) | Parabaeus papei Buhl |
|
||
| – | Metasomal horn distinct, two-thirds of dorsal length of mesoscutum (a, b), excavation encompasses c. a quarter of mesoscutal length (a, b) | Parabaeus brevicornis Buhl |
|
||
| 9 | Posterior margin of mesosoma broadly pointed in dorsal view, with short longitudinal carinae present along posterior border (A); in lateral view mesosoma gently humped one-third of distance from posterior margin (B) | Parabaeus africanus Austin |
|
||
| – | Posterior margin of mesosoma rounded in dorsal view, with narrow smooth posterior border containing scattered short setae in pits (a); in lateral view mesosoma without dorsal hump (b) | Parabaeus ruficornis Kieffer |

|
||
Parabaeus abyssus
Parabaeus abyssus
Austin, 1990: 649, 650 (original description, species key, illustrated);
Material examined
Holotype
: Australia • ♀; Western Australia, Perth, Kings Park; 1952; G. Bornemissza (

Paratype
: Australia • ♀; Western Australia, Walpole-Nornalup Nat. Pk; 34°59'S, 116045'E; 17–21.i.1987; J. S. Noyes (
Distribution
Australia (
Parabaeus africanus
Parabaeus africanus
Austin, 1990: 649, 655 (original description, species key, illustrated);
Material examined
Holotype
: Kenya • ♂; Mt Kulal; 2134 m; April 1980; D. Levin (
Distribution
Malawi (
Parabaeus armadillus
Parabaeus armadillus
Austin, 1990: 649, 658 (original description, species key, illustrated);
Material examined
Holotype
: South Africa • ♀; Port St Johns, Pondoland; Dec. 1923; R. E. Turner; Brit. Mus. 1924-97 (

Paratypes
: South Africa • 5♀♀; 2 unknown sex: same data as holotype, but with different dates (Nov. 1923–25.ii.1924) • 3♀♀; 1 unknown sex (

Additional material
SOUTH AFRICA • 1♀, 1♂; Kwazulu-Natal, Umtamvuna Nature Reserve; 31°03.506'S 30°10.392'E, 160m; 15-16.xi.2000; S. van Noort; Malaise trap; KW00-M74; Coastal Forest; SAM-HYM-P031787A; SAM-HYM-P031787B (

Comments
There is the possibility that the additional material cited above may include two similar looking species. There is some variation present within the available material with regard to colouration, dimensions of the mesosoma, and degree of striation on the metasoma. Whether this is intra-specific variation, or indicative of the presence of additional species, requires a focused morphologically assessment, ideally with the additional aid of barcoding tools.

Distribution
South Africa (
Parabaeus austini
Parabaeus austini Buhl, 2011: 95 (original description, illustrated).
Material examined
Holotype
: Tanzania • ♀; Tanga, Lushoto Dist., Mazumbai For. Res.; 1370–1435 m; 4.xii.1995 (
Distribution
Tanzania (
Parabaeus brevicornis
Parabaeus brevicornis Buhl, 2011: 95 (original description, illustrated).
Material examined
Holotype
: Tanzania • ♀; Tanga, Muheza Dist., Kwamgumi For. Res.; 170–220 m; 18.vii.1995 (
Distribution
Tanzania (
Parabaeus nasutus , sp. nov.
Material examined
Holotype
: South Africa • ♀; Northern Cape, Swaarweerberg, Vredehoek Farm; 1613 m; 32°26.387'S, 20°34.501'E; 29 July–30 September 2010; S. van Noort; Yellow pan trap; Roggeveld Shale Renosterveld; SWA09-SUC1-Y05; SAM-HYM-P040757 (

Paratypes
: South Africa • 3♂♂, data as for holotype, except for 31 March–29 July 2010; SWA09-SUC1-Y04; SAM-HYM-P040756a-c (

Parabaeus nasutus van Noort sp. nov. female holotype (
Excluded from type material
South Africa • 1♀; Western Cape, Gamkaberg Nature Reserve; 33°39.941'S, 21°53.505'E; 315 m; 19 Feb–30 Mar 2010; S. van Noort; Yellow pan trap; Gamka Thicket; GB09-SUC1-Y28; SAM-HYM-P093813 (
Description
Female body length: 0.84 mm. Colour of head, metasoma, antennae and fore and mid legs brown; mesosoma and hind legs yellow-brown.
Head as wide as long. Much wider than mesosoma, fractionally narrower than width of metasoma; in dorsal view moderately transverse; clypeus produced into flattened volcano-shaped, nasute-like process with central fovea ringed by a carina; malar sulcus present; frons convexly rounded; occiput vertical, mostly hidden by mesosoma; occipital carina not visible dorsally, not reaching to posterior margins of eyes or lateral ocelli; lateral ocelli connected by hyperoccipital carina forming a sharp dorsal delimitation between occiput and vertex; ocelli forming an obtuse triangle, POL>LOL; in anterior view frons 0.6× width of head; subocular carina absent; gena wide; head covered with coriaceous sculpturing; antennal segments short and robust, clava 2.5× as long as wide.
Mesosoma. Robust, 0.8× width of metasoma, as long as wide; in dorsal view pronotum only visible as narrow strip around anterior margin, pronotal collar with posteriorly orientated, raised, medial bifurcated projection; mesoscutum convexly rounded, posterior margin strongly elevated, with scutellum forming dorsal plateau defined by darker, toothed ellipsoidal carina; posterior face abruptly declivitous; in lateral view pronotal spiracle seen as a small toothed bump at posterodorsal corner of pronotum; tegula absent; mesopleural carina absent; mesopleural carina present; dorsal mesosoma imbricate with associated posteriorly facing setae; pronotum imbricate in dorsal half, smooth with longitudinal striations in ventral half of lateral face; mesopleuron smooth dorsally with longitudinal striations in ventral half; pronotum and mesopleuron fused; metapleuron and dorsolateral propodeum densely covered with fine setae.
Metasoma. In dorsal view oval in shape; anterior margin broad; T1 very narrow, densely setose laterally, base of metasoma without obvious foveate pits (although there are possibly two pits indicated by depressions that are obscured by setae); tergite 1 with bifurcate projecting medial plate; T2 composing virtually all dorsal metasoma; T3–T6 very narrow, strip-like, only seen in posterior view; anterior T2 faintly longitudinally coriaceous, smooth in posterior half, whole surface sparsely setose.
Male. As in females, except for clava, which is more elongate.
Diagnosis
Parabaeus nasutus has the following unique morphological apomorphies: elevation of posterior section of mesoscutum and scutellum into a medial projection, which dorsally has a transversely ellipsoidal plateau formed by the scutellum with a 90 degree drop-off posteriorly; base of metasoma without obvious foveolate pits (although there are possibly two pits indicated by depressions that are obscured by setae); T1 with bifurcate projecting medial plate; pronotal collar with posteriorly projecting medial bifurcating raised plate; clypeus produced into dorso-ventrally compressed, volcano-shaped, nasute-like process with a central apical fovea ringed by a carina; occipital carina not visible dorsally. Although P. nasutus has a number of derived diagnostic characters, from a ground-plan perspective the species is morphologically similar to P. peckorum sharing the same squamate sculpturing with scattered posteriorly projecting setae on the dorsal mesosoma, and dense setose patches on the metapleuron, dorsolateral propodeum and T1. Colour is, however, different from P. peckorum, which has a dark brown to black body with lighter brown antennae and legs, and dense white pubescence at the mesosomal-metasomal boundary.
Etymology
Named for the exceptional clypeal modification into a nasute-like process. Latin adjective.
Distribution
South Africa.
Comments
We suspect that the central fovea ringed by a carina that is terminally situated on the clypeal nasute-like process is olfactory in nature, potentially containing chemo-sensillae that may be involved in host location, although males also possess this adaptation, so possibly it is involved in mate recognition. It is likely that the species lives in the leaf-litter habitat and probably attacks insect or arachnid eggs.
The single female from Gamkaberg Nature Reserve is uniformly orange-yellow, has a smoother mesopleuron, and weaker clypeal and posterior mesosomal protrusions. Overall, the surface sculpturing is also weaker. The specimen is smaller than the type series specimens and these differences may simply be related to the reduced size. There is, however, the possibility that this specimen represents a second closely related, undescribed species, but until further specimens are acquired to assess the degree of intraspecific variation this specimen is considered to belong to P. nasutus, but it is excluded from the type material.
Parabaeus papei
Parabaeus papei Buhl, 2011: 96 (original description, illustrated).
Material examined
Holotype
: Tanzania • ♀; Tanga, Lushoto Dist., Mazumbai For. Res.; 1370–1435; 8.xii.1995 (
Distribution
Tanzania (
Parabaeus peckorum
Parabaeus peckorum
Austin, 1990: 657 (original description, species key, illustrated);
Material examined
Holotype : South Africa • ♀; Natal, 75 km WSW Estcourt, Cathedral Peaks, Rainbow Gorge, podocarp forest; 1500 m; sweeping; 17.xii.1979; S. and J. Peck (SANC).
Paratypes
: South Africa • 3♀♀; same data as holotype; 2♀♀ (
Distribution
South Africa (
Parabaeus quasimodus
Parabaeus quasimodus
Austin, 1990: 655 (original description, species key, illustrated);
Material examined
Holotype
: Kenya • ♀; Mt Kulal; 2134 m; April 1980; D. Levin (
Distribution
Kenya (
Parabaeus ruficornis
Parabaeus ruficornis
Kieffer, 1910: 294 (original description);
Material examined
Holotype
: Seychelles • ♀; Mahe; Percy Sladen Trust Expedition; 1913-170; ‘08-9 [= August-September?]; figured specimen; B.M. TYPE HYM 9.399 (

Distribution
Seychelles (
Pulchrisolia
Pulchrisolia
Szabó, 1959: 395 (original description. Type: Pulchrisolia maculata Szabó, by monotypy and original designation);
Diagnosis
Fore wing with very short, tubular R vein terminating in a knob and at least some microtrichia of the fore and hind wings in the form of short, scale-like pegs; distinctive colour of the adult (most species are yellow, orange, red, or a combination thereof); frontal ledge present on the lower frons in all but one species; interantennal process present, bilobed in most species; and tract of dense setae on the metatibia.

Species richness
Pulchrisolia ankremos Lahey, 2019 (Ghana, Ivory Coast).
Pulchrisolia asantesana van Noort & Lahey, 2019 (South Africa).
Pulchrisolia diehoekensis van Noort & Lahey, 2019 (South Africa).
Pulchrisolia ellieae Lahey, 2019 (Madagascar).
Pulchrisolia maculata Szabó, 1959 (Kenya, Tanzania).
Pulchrisolia nephelae Lahey, 2019 (Benin, Burkina Faso, Gambia, Ivory Coast, Mali, Nigeria).
Pulchrisolia robynae van Noort & Lahey, 2019 (South Africa) (Fig.
Pulchrisolia sanbornei Lahey & Masner, 2019 (South Africa).
Pulchrisolia teras Lahey, 2019 (Madagascar).
Pulchrisolia valerieae Polaszek & Lahey, 2019 (Zambia).
Distribution
Endemic to the Afrotropical region: Benin, Burkina Faso, Gambia, Ghana, Ivory Coast, Kenya, Madagascar, Mali, Mozambique, Nigeria, South Africa, Tanzania, Zambia (
Biology
Unknown.
Key to species of Pulchrisolia
Pulchrisolia ankremos
Pulchrisolia ankremos Lahey, 2019, in Lahey et al. 2019: 44, 49 (keyed, description).
Material examined
Holotype
: Ghana • ♀; Ashanti Reg., Bobiri Forest Reserve; 06°42'N, 01°20'W; II-2002; flight intercept trap; C. Carlton & O. Frimpong; OSUC 20666426 (
Distribution
Ghana, Ivory Coast (
Pulchrisolia asantesana
Pulchrisolia asantesana van Noort & Lahey, 2019, in Lahey et al. 2019: 45, 49, 50, 51, 52 (keyed, description).
Material examined
Holotype
: South Africa • ♀; Eastern Cape Prov., Asante Sana Game Reserve, Zuurkloof; 1621 m; 32°16.011'S, 25°00.244'E; 23.X.2010; pitfall trap; J. Midgley; T2S3d; Camdeboo Escarpment Thicket, tall grass stands, scattered oubos shrubs; SAM-HYM-P046628a (
Distribution
South Africa (
Pulchrisolia diehoekensis
Pulchrisolia diehoekensis van Noort & Lahey, 2019, in Lahey et al. 2019: 45, 52 (keyed, description).
Material examined
Holotype
: South Africa • ♀; Eastern Cape, Winterberg, The Hoek Farm; 1879 m; 32°21.260'S, 26°23.001'E; 9 April –26 July 2010; S. van Noort; yellow pan trap; Amathole Mistbelt Grassland; WTB09-GRA1-Y04; SAM-HYM-P038987 (
Distribution
South Africa (
Pulchrisolia ellieae
Pulchrisolia ellieae Lahey, 2019, in Lahey et al. 2019: 44, 53, 54, 55 (keyed, description).
Material examined
Holotype : Madagascar • ♀; Toliara Auto. Prov., 60 km NE Morondava, Beroboka Avaratra; 18.V–23.V.1983; J. S. Noyes & M. C. Day; OSUC 20666430 (NMHUK).
Distribution
Madagascar (
Pulchrisolia maculata
Pulchrisolia maculata
Szabó, 1959: 396 (original description);
Sceliotrachelus maculatus
(Szabó, 1959):
Material examined
Holotype
: Tanzania • ♀; Mara Reg., Shirati, V-1909; Katona; Hym. Typ. No. 9583 Mus. Budapest. (
Distribution
Kenya, Tanzania (
Pulchrisolia nephelae
Pulchrisolia nephelae Lahey, 2019, in Lahey et al. 2019: 44, 57, 58, 59 (keyed, description).
Material examined
Holotype
: Mali • ♀; Koulikoro Reg., Mourdiah; 25.VIII–5.IX.1986; Malaise trap; M. Matthews; OSUC 20666433 (
Distribution
Benin, Burkina Faso, Gambia, Ivory Coast, Mali, Nigeria (
Pulchrisolia robynae
Pulchrisolia robynae van Noort & Lahey, 2019, in Lahey et al. 2019: 45, 59, 60, 61 (keyed, description).
Material examined
Holotype
: South Africa • ♀; Eastern Cape Prov., Marais Hoop Farm, 25.6 km (254°) W Kirkwood; 33°32.635'S, 25°13.678'E; H. G. Robertson & R. Tourle; valley bushveld (goat trashed); pitfall trap, VB01-R4T-P06; SAM-HYM-P031619 (
Distribution
South Africa (
Pulchrisolia sanbornei
Pulchrisolia sanbornei Lahey & Masner, 2019, in Lahey et al. 2019: 45, 61 (keyed, description).
Material examined
Holotype
: South Africa • ♀; Limpopo Prov., 15 km E Klaserie, Guernsey Farm; 19.XII–31.XII.1985, pan trap, M. Sanborne, OSUC 20666387 (
Distribution
South Africa (
Pulchrisolia teras
Pulchrisolia teras Lahey, 2019, in Lahey et al. 2019: 44, 62, 63, 64, 65, 66, 67 (keyed, description).
Material examined
Holotype : Madagascar • ♀; Toliara Auto. Prov., Andohahela National Park, 36.1 km (308°) NW Tolagnaro, 1.7 km (61°) ENE Tsimelahy, Ambohibory Forest; 300 m; 24°55'48"S, 46°38'44"E; BLF4915; 16.I–20.I.2002; pitfall trap; Fisher, Griswold et al.; CASENT 2043862 (CASC).
Distribution
Madagascar (
Pulchrisolia valerieae
Pulchrisolia valerieae Polaszek & Lahey, 2019, in Lahey et al. 2019: 44, 67, 68, 69, 70 (keyed, description).
Material examined
Holotype
: Zambia • ♂; Lukwakwa, open Dambo; 12°39'40"S, 24°26'13"E; 1147 m; 4–8.ix.13; Yellow Pan; Smith, Takano and Oram; NHMUK010823075, type number 9.1020 (
Distribution
Zambia (
Sceliotrachelus
Sceliotrachelus
Brues, 1908: 13 (original description; type: Sceliotrachelus braunsi Brues, 1908, by monotypy and original designation).
Pulchrisolia
Szabó, 1959: 395 (original description. Type: Pulchrisolia maculata Szabó, by monotypy and original designation);
Generic redescription to accommodate two new additional species
Colour of head and mesosoma black; metasoma black to light brown or orange-brown; scape, pedicel, flagellar segments in female, legs orange-brown; club and flagellar segments in male black; wings either infuscate or dark with white areas.
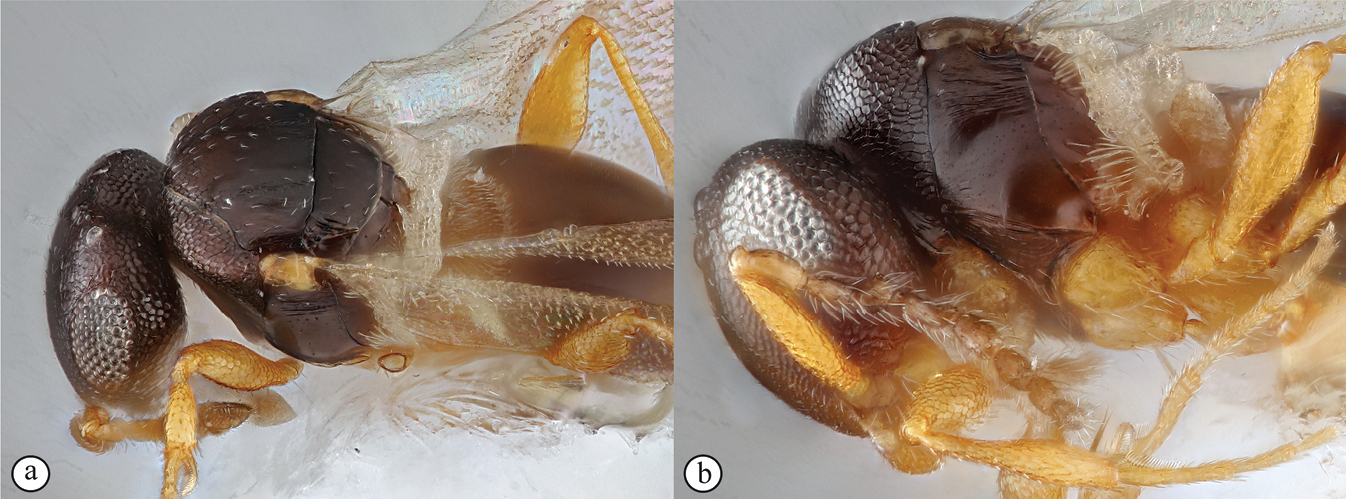
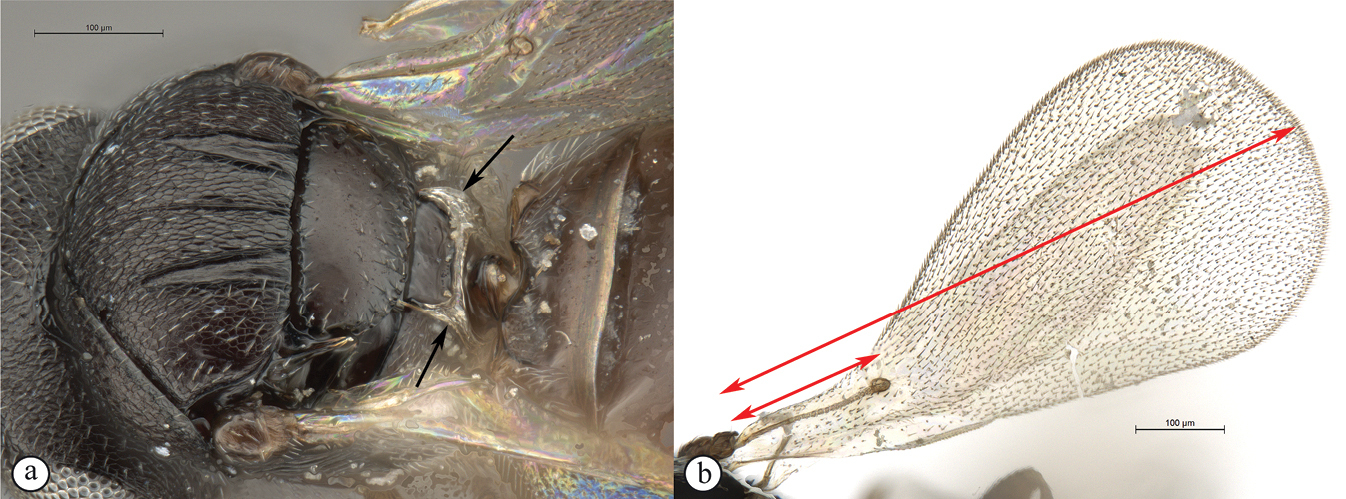
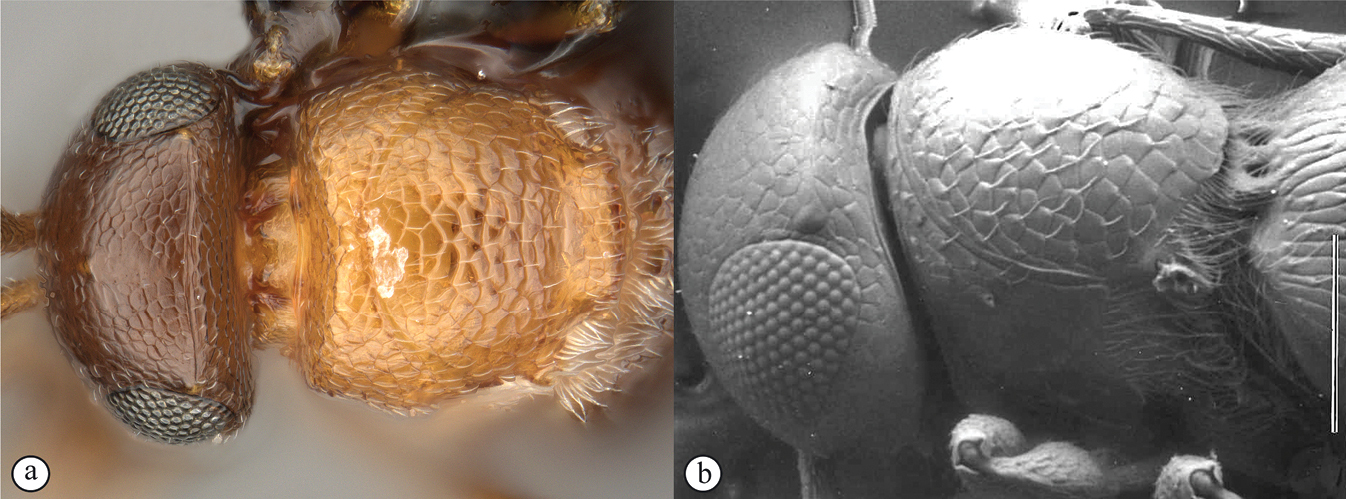
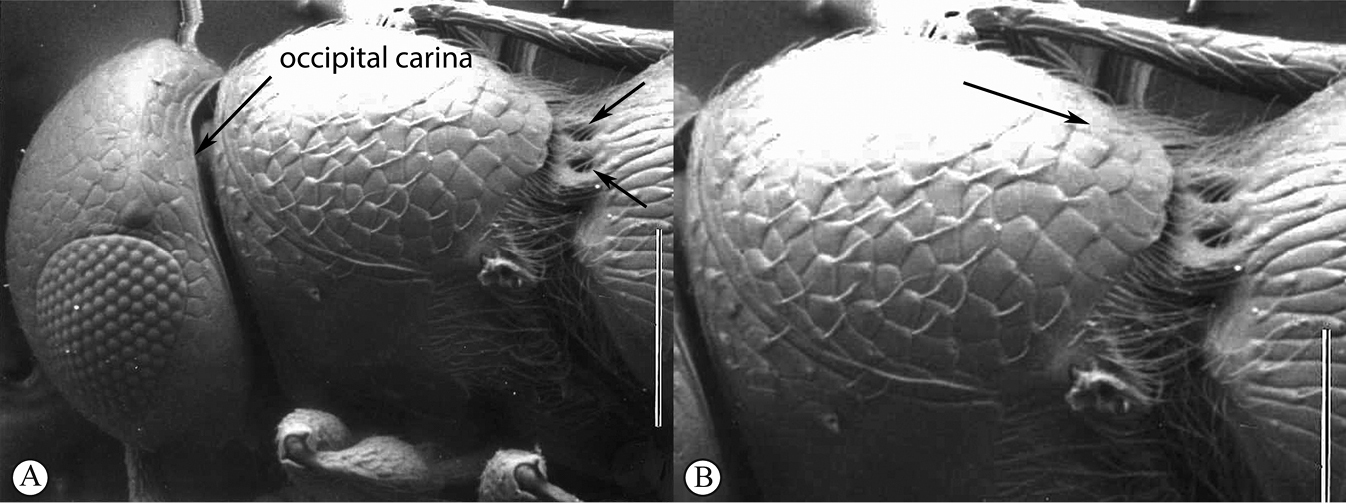
Head wider than long, narrowing towards clypeus; frontal ledge absent; antenna 10-merous; clava subcompact, terminal segment tapering, 3-merous; arrangement of setae on ventral surface of each clavomere forming a chevron-shaped area with the posterior-most papillary sensillum at its point; male antennae filiform; toruli in close apposition, separated by less than one torular diameter, positioned on frontal protrusion close to mandibles, situated well below compound eyes; inter-antennal process present, acuminate distally; clypeus smooth, with convex margin; mandibles bidentate; frons micro-reticulate, finely punctate in dorsal half and along inner orbits, with minute setae medially; malar sulcus absent, malar space three-fifths of eye height; facial and malar striae absent; hyperoccipital carina present on dorsal margin of vertex, anterior profile with two raised areas corresponding with lateral ocelli; vertex anterior to hyperoccipital carina finely punctate, with minute setae, posterior of carina micro-reticulate; lateral ocelli positioned posterior of hyperoccipital carina, separated from inner margin of compound eye by more than 5 ocellar diameters; occiput micro-reticulate; occipital carina present, with or without occipital pit; occipital carina ventrally reaches, or approaches the anterior articulation of the mandible.
Mesosoma. Pronotal shoulder sharply angled transversely, with pronotal carina present posteriorly; pronotum transverse, weakly to strongly triangular with strong medial longitudinal sulcus (possibly representing convergence of two epomia); shoulders weak to strong, micro-reticulate to polished; pronotal cervical sulcus with depressions at ventral and dorsal apices, dorsal depression setose; admedian depressions present, widely spaced, longitudinally offset from deep posterior notaular grooves; mesoscutellar disc flanked by parallel longitudinal grooves, which fuse with the trans-axillar carina; axillar carinae present or absent; axillae moderately to strongly excavated; sculpture of mesoscutellum micro-reticulate to polished; mesopleuron polished, may be dorsally transversely ridged, or longitudinally compressed, much higher than long; transepisternal line absent; mesopleural carina strong, may form a flange posteriorly; foamy structures present posteriorly on metapleuron, concealing metapleural carina; metapleural pit present; submarginal vein of fore wing absent or spectral; marginal cilia of fore wing absent; wing microtrichia normal, or strong and needle-like; hind wing may have a strong thickened marginal vein.
Metasoma. Ovate, sessile with indistinct lateral carina; T1 transverse in dorsal view; T2 large, as long as wide, comprising more than half to 4/5ths of metasomal length; foamy structures present anterolaterally on T1 and on S1; posterior margin of T1 with fringe of long setae; anterior margin of T2 with transverse furrow covered by elongate setae of posterior margin of T1, containing minute setae that often accumulate a white exudate; scattered long setae present on S2; tibial spur formula 1-2-2; protibial spur with comb of setae.
Diagnosis
Sceliotrachelus is recognizable by the median longitudinal sulcus on the pronotum and the presence of long setae on sternite 2. These two characters separate the genus from all other sceliotrachelines. Additional diagnostic characters are: the malar sulcus with tract of long, straight setae; hyperoccipital carina present with the lateral ocelli positioned posterior to its margin; widely spaced admedian depressions on the mesoscutum; notauli present; transaxillar and axillular carinae fused; axillar area as wide as or wider than mesoscutellum; transepisternal line absent; metapleuron with lateral projection in ventral half; ventral surface of coxae, trochanters, and portion of femur and S2 with long setae; scrobe present on dorsal surface of hind femur; foamy structures present on propodeum, metapleuron, T1, and S1; anterior margin of T2 with a transverse, setose furrow; submarginal vein of fore wing absent.
Sceliotrachelus shares morphological affinities with Afrisolia, Isolia and Pulchrisolia, together forming the Isolia-cluster, which is defined by the combed fore tibial spur. Additional putative synapomorphic characters centre on the foamy structures present on the propodeum in these four genera (
Species richness
Sceliotrachelus braunsi Brues, 1908 (South Africa) (Figs
Sceliotrachelus karooensis van Noort, sp. nov. (South Africa) (Figs
Sceliotrachelus midgleyi van Noort, sp. nov. (South Africa) (Figs
Distribution
The genus Sceliotrachelus is, as far as known, confined to the Eastern and Western Cape Provinces of South Africa (Fig.
Biology
Unknown, but likely to be parasitoids of arthropods living in the leaf litter habitat (see discussion).
Key to species of Sceliotrachelus
| 1 | Pronotum and occiput glabrous (A); occipital pit present (A); acetabular carina (ac) and mesopleural epicoxal sulci (mes) not converging, fore and mesocoxae separated by more than one fore coxal width (B); fore wing of normal shape, twice as long as wide (C) | Sceliotrachelus karooensis van Noort, sp. nov. |

|
||
| – | Pronotum and occiput setose (a); occipital pit absent (a); acetabular carina (ac) and mesopleural epicoxal sulci (mes) converging, fore and mesocoxae separated by less than one fore coxal width (b); fore wing narrow and elongate, at least 3× longer than wide, anterior and posterior margins subparallel (c) | 2 |

|
||
| 2 | Genal and pronotal rugae present (A); mesoscutellum compressed, narrow, 2.5× wider than long (B); mesoscutellar disc medially with broad, shallow, longitudinal excavation, axillar carinae weakly raised (B); costal margin of hind wing with thick band of black sclerotization that runs nearly the entire length of the wing (B); fore wing extending beyond posterior margin of T2 | Sceliotrachelus braunsi Brues |

|
||
| – | Genal and pronotal rugae absent (a); mesoscutellum broader, twice as wide as long (a); mesoscutellar disc square, medially and longitudinally evenly raised into a hump, laterally defined by strongly raised longitudinal axillar carinae (b); costal margin of hind wing not sclerotized; fore wing not reaching posterior margin of T2 (c) | Sceliotrachelus midgleyi van Noort, sp. nov. |

|
||
Sceliotrachelus braunsi
Sceliotrachelus braunsi
Brues, 1908: 13 (original description);
Material examined
Holotype
: South Africa • ♂; Eastern Cape, Algoa Bay, Cape Colony; 10 November 1896; H. Brauns; Sceliotrachelus braunsi Brues (

Sceliotrachelus braunsi holotype male (
Paratype
: South Africa • ♂; same data as holotype (

Additional material examined
South Africa • ♀; Eastern Cape: 1♀; 1♂ Schilpad Laagte Farm, (14.7 km 229°SW Kirkwood); 33°31.104'S, 25°22.353'E; 9–16 Feb 2001; HG Robertson and R Tourle; Pitfall; VB01-A2T-P02; Valley Bushveld (goat trashed) [Sundays Thicket]; SAM-HYM-P030896 (
Description
Female body length: 2 mm; male body length: 2 mm. Colour of head and mesosoma black; metasoma light brown with lateral tergites below lateral keel orange-brown; scape, pedicel, flagellar segments in female, legs orange-brown; flagellar segments in male black; foamy structures white; wings with dark brown and white patches.

Head 1.15× wider than long, strongly narrowing towards toruli and clypeus; eye height equal to inter-ocular distance; malar space 0.6× eye length; antennal length ratios (female): scape 13, pedicel 3, funicle 5, clava 10; male antennae filiform: scape 13, pedicel 3, flagellum 30; LOL:OOL:POL (2.5:5.5:5.5); occipital carina present, without occipital pit with numerous long white setae.


Metasoma. T1 transverse in dorsal view with dense row of setae on posterior margin overlapping T2; T2 large, as long as wide, 0.7× metasomal length, with anterior transverse furrow c. equivalent to length of T1, centrally situated, encompassing two-thirds of anterior tergite width; T2 anteriomedially polished grading into posterior micro-reticulate area, bounded anterolaterally by more strigate area.

Male as in female, except for antennal configuration: eight funicular segments, first as long as second and third combined; second to seventh subequal in length, 2× longer than wide; ultimate segment longer than penultimate segment.
Diagnosis
The shape and colour pattern of the fore wings immediately distinguish this species from the other two species, which either have a much more brachypterous or normal fore wing shape. The hind wing costal margin has a thick band of black sclerotization that runs nearly the entire length of the wing, which is absent in the other two species. Strong genal and pronotal rugae are present, absent in the other two species. The mesosoma is the most longitudinally compressed of the three species with the pronotum, mesoscutum and scutellum all extremely transverse and of equal length. Strong white setae are present on the occiput and pronotum. The mesoscutum is posteriorly strongly raised in lateral view; mesoscutum compressed, narrow 2.5× wider than long, without raised carinae; pronotal shoulders taper to point; wings slightly shortened, extending just beyond posterior margin of second tergite, 3× longer than wide.

Etymology
Named by Brues after the collector of the two type specimens, Dr Hans Heinrich Justus Carl Ernst Brauns, a medical doctor who practiced in Willomore in the Eastern Cape. The Brauns collection of Apocrita Hymenoptera was purchased by the Transvaal Museum (now Ditsong Museums of South Africa) for £1500 (

Sceliotrachelus braunsi female SEM photos (
Distribution and habitat association
(Fig.
Gamtoos Thicket (coastal basin of the Gamtoos River Valley, south of the Baviaanskloof Mountains and along some smaller river valleys such as that of the Kromme River; also found north of the Baviaanskloof Mountains in more xeric conditions on some low ridges south and southeast of Steytlerville; altitude 0–700 m).
Sundays Noorsveld (mostly north of the Klein Winterhoek Mountains, centred around Waterford and the Darlington Dam and a smaller area from Jansenville westwards; also some patches south of this mountain range west of Kirkwood in the Sundays River Valley; altitude 100–600 m).
Sundays Thicket (from the surrounds of Uitenhage and the northern edge of Port Elizabeth into the lower Sundays River Valley to east of Colchester and northwards to the base of the Zuurberg Mountains and stretching westwards north of the Groot Winterhoek Mountains to roughly the Kleinpoort longitude; also an extensive area north of the Klein Winterhoek Mountains including much of the Jansenville District and parts of the far-southern Pearston District and far-western Somerset East District; altitude 0–800 m).

The current distribution is likely to be an artefact of under-sampling and the species is expected to be more widespread in the Eastern Cape (Fig.
Comments
The female specimen (Fig.
Sceliotrachelus karooensis , sp. nov.
Material examined
Holotype
: South Africa • ♀; Western Cape, Anysberg Nature Reserve, 5.8 km west of Vrede; 785 m; 33°28.658'S, 20°31.572'E; 8 Dec 2014–8 Jan 2015; S. van Noort; yellow pan trap; Succulent Karoo; ANY14-SUC1-Y19; SAM-HYM-P086434 (

Paratypes
: South Africa • 1♀; same data as holotype, except SAM-HYM-P086435 (

Sceliotrachelus karooensis van Noort, sp. nov. female holotype (
Description
Female body length: 2.4 mm; male body length: 2.3 mm. Colour of head and mesosoma black; metasoma light brown anterolaterally, with central tergites and sternites orange-brown; scape, pedicel, flagellar segments in female, legs orange-brown; flagellar segments in male black; foamy structures pale yellow; wings infuscate.

Head 1.27× wider than long, strongly narrowing towards toruli and clypeus; eye height 0.9× inter-ocular distance; malar space 0.55× eye height; antennal length ratios (female): scape 16, pedicel 3, funicle 8, clava 12; male antennae filiform: scape 23, pedicel 5, flagellum 60; LOL:OOL:POL (4:7:7); hyper-occipital carina present, collared, with occipital pit, glabrous.

Sceliotrachelus karooensis van Noort, sp. nov. male paratype (
Mesoscutum. Sculpture of mesosoma polished; pronotum transverse, weakly delta-shaped with medial longitudinal sulcus, glabrous; shoulders rounded, flat in lateral view; mesoscutum short 2.4× wider than long medially; 3× medial length of pronotum, equivalent to scutellar length; anterior admedian depressions short, deep grooves, not overlapping longitudinally with posterior shallow notauli, which are widely spaced; weak median mesoscutal line present; notauli align with grooves on lateral margins of mesoscutellar disc, which are continuous with the scutoscutellar sulcus, itself widened; axillar carinae present; axillae weakly and evenly excavated; posterior margin of mesoscutellum not raised in lateral view; propodeum with foamy structures extending to T1; mesopleuron polished, dorsally with c. ten incomplete transverse ridges; not longitudinally compressed, 1.4× higher than long; mesopleural carina strong, curled dorsally; foamy structures present posteriorly on metapleuron, concealing metapleural carina, ventrally with dense patch of yellow setae; fore wing of normal shape, 2.25× longer than wide, reaching to end of metasoma, almost glabrous with scattered microtrichiae in basal quarter, dense microtrichiae present over distal three-quarters; submarginal vein of fore wing present, but spectral and not reaching margin; hind wing with submarginal vein absent, except for basal remnant that has stout setae, anterior margin straight, slightly humped at hamulus, which comprises strong, curved setae, narrow and almost glabrous with scattered microtrichiae on basal half, dense microtrichiae present over distal half; long setae present on basal half of posterior wing margin.

Sceliotrachelus karooensis van Noort, sp. nov. A–C female paratype (
Metasoma. T1 transverse in dorsal view with dense row of setae on posterior margin overlapping T2; T2 large, wider than long, 0.63× metasomal length, with anterior transverse furrow 0.5× length of T1, centrally situated, encompassing central three-quarters of anterior tergite width; T2 centrally polished grading into lateral micro-reticulate area, bounded anterolaterally by more strigate area.

Sceliotrachelus karooensis van Noort, sp. nov. female paratype SEM photos (
Male as in female, except for antennal configuration: 8 funicular segments, first longer than second; second to seventh subequal in length, 3.5× longer than wide; ultimate segment longer than penultimate segment.
Diagnosis
This species is immediately identifiable by the appearance of the fore wings which are of normal shape, and uniformly fuscous, without dark patches or strong microtrichiae, in contrast to the elongate and chromatically modified fore wings in the other two species. An additional diagnostic character is the presence of an occipital pit, which is absent in the other two species. The mesosoma is correspondingly less longitudinally compressed, with the mesocutum and mesopleuron being more normal in proportions. The pronotum and occiput are glabrous without the long setae present in the other two species. The acetabular and mesopleural epicoxal sulci do not converge as in the other two species, and, correspondingly, the fore and mesocoxae are separated by more than one fore coxal width (separated by less than the fore coxal width in other two species).
Etymology
Named after the Karoo area wherein both main collection localities fall. Noun in apposition.
Distribution and habitat association
(Fig.
Western Little Karoo (the unit covers most of the western basin of the Little Karoo from the confluence of the Groot and Gouritz Rivers in the west as far as Anysberg by surrounding this mountain range and also extending along the northern flanks of the Klein Swartberg; two larger patches of the Western Little Karoo are found immediately to the east and south of Touws River, and one small isolated patch fringes the Langeberg Mountains in the Montagu area; altitude 160–1060 m (most of the area at 300–860 m).
South Rooiberg Sandstone Fynbos (southern slopes of the mountains of Rooiberg, Gamka and the Amalienstein Ridge-Sandberg-Bakenskop range; altitude 350–1490 m on the summit of Rooiberg).
The current distribution is likely to be an artefact of under-sampling and the species is expected to be more widespread in the Western Cape.
Sceliotrachelus midgleyi , sp. nov.
Material examined
Holotype
: South Africa • ♀; Eastern Cape, Asante Sana Game Reserve, Zuurkloof; 32°17.092S, 25°00.521E; 1204 m; 18–25 Nov 2009; J. Midgley; T2S1c; Pitfall trap; Camdeboo Escarpment Thicket; Acacia thicket; next to river bed with larger trees; SAM-HYM-P046650 (

Paratypes
: South Africa • 3♀♀, 1♂; Eastern Cape, Asante Sana Game Reserve, buttress between Waterkloof and Zuurkloof; 32°16.932S, 24°58.795E; 1401 m; 18–25 Nov 2009; J. Midgley; T1S1e; Pitfall trap; Camdeboo Escarpment Thicket, Open habitat, low shrubs and mat forming wild carnation, scattered grass; SAM-HYM-P036025A-D (

Description
Female body length: 2.26 mm; male body length: 2 mm. Colour of head and mesosoma black; metasoma dark brown to black, with lighter brown areas anteriolaterally; scape, pedicel, flagellar segments in female, legs orange-brown; flagellar segments in male dark brown; foamy structures pale yellow; wings yellow-white with broad darker band in apical half.

Head 1.27× wider than long, strongly narrowing towards toruli and clypeus; eye height 0.9× inter-ocular distance; malar space 0.55× eye length; antennal length ratios (female): scape 14, pedicel 2.5, funicle 5.5, clava 10; male antennae filiform: scape 15, pedicel 4, flagellum 35; LOL:OOL:POL (4:7:7); hyperoccipital carina present, collared, without occipital pit, long setae present.

Sceliotrachelus midgleyi van Noort, sp. nov. A, B female paratype (
Mesoscutum. Sculpture of mesosoma polished; pronotum transverse, strongly delta-shaped with medial longitudinal sulcus, with anteriorly projecting neck, covered in scattered long brown setae; shoulders angular, flat in lateral view; mesoscutum short 2.3× wider than long medially; 1.3× medial length of pronotum and equivalent to scutellar length; admedian depressions short, deep grooves, not overlapping longitudinally with posterior narrow and more shallow notauli, which are widely spaced; median mesoscutal line present; notaular excavations align with two deep and long parallel excavations of the axillular carina, which meet with the scutoscutellar sulcus, itself not widened; mesoscutellar disc medially and longitudinally evenly raised into hump; axillar carinae present and strongly raised; axillae weakly and evenly excavated; posterior margin of mesoscutellum not raised in lateral view; propodeum with foamy structures extending to T1; mesopleuron polished, dorsally with two complete transverse ridges; not longitudinally compressed, 1.9× higher than long; mesopleural carina strong, curled dorsally; foamy structures present posteriorly on metapleuron, concealing metapleural carina, metapleuron ventrally with dense patch of white setae; fore wing brachypterous, not extending beyond posterior margin of T2, narrow > 4× longer than wide, curved forwards in boomerang-shape; submarginal vein of fore wing absent except at base, narrow and almost glabrous with scattered microtrichiae for basal third, dense microtrichiae present over distal two-thirds; hind wing with marginal vein absent, except for basal remnant, anterior margin angled at hamulus, narrow and almost glabrous with scattered microtrichiae for basal half, dense microtrichiae present over distal half.

Metasoma. T1 transverse in dorsal view with dense row of setae on posterior margin overlapping T2; T2 large, as long as wide, 0.7× metasomal length, with anterior transverse furrow 0.5× length of T1, centrally situated, encompassing central three-quarters of anterior tergite width; T2 anteromedially polished grading into lateral strigulate areas, and posteriorly aerolate-rugulose.

Sceliotrachelus midgleyi van Noort, sp. nov. female paratype SEM photos (
Male as in female, except for antennal configuration: eight funicular segments, first 1.3× as long as second and third combined; second to seventh subequal in length, 2× longer than wide; ultimate segment longer than penultimate segment.
Diagnosis
This species is immediately distinguishable by the brachypterous wings, which have a unique colour pattern, are extremely narrow, 4× longer than wide, and do not extend beyond the posterior margin of tergite 2. It shares the lack of the occipital pit, presence of long setae on the pronotum, and presence of strong microtrichiae with S. braunsi, but does not have rugae present on the genae and pronotum as in S. braunsi; has a wider mesoscutum (twice as wide as long, as opposed to 2.5× in S. braunsi); a distinctive mesoscutellar disc which is square in shape and medially and longitudinally evenly raised into a hump, laterally defined by strongly raised longitudinal axillar carinae; and the costal margin of hind wing is not sclerotized as in S. braunsi.

Etymology
Named in honour of John Midgley, who first collected specimens of the new species as part of his PhD project. Noun in the genitive case.
Distribution and habitat association
(Fig.
Camdebo Escarpment Thicket (south-sloping face of the Great Escarpment, forming an arc from Bruintjieshoogte in the east via the Coetzeeberg Mountains and Graaff-Reinet (including Spandaukop and the isolated Rooiberg) to Kamdebooberg and Aberdeen in the west; altitude varies from 570–1600 m, with most of the area between 700–1200 m).
The current distribution is likely to be an artefact of under-sampling and the species is expected to be more widespread in the Eastern Cape, but possibly restricted to higher altitudes in Camdebo Escarpment Thicket.
Discussion
The circumscription and phylogenetic affinities of the subfamily Sceliotrachelinae are not yet fully resolved. The extreme adaptive morphology that has likely been driven by selection emanating from life history strategy clouds assessment of the evolutionary relationships based on morphology. Evolutionary selection can readily result in convergence when the same ecological pressures are acting on distinct evolutionary lineages. Historical molecular analysis (
Based on current specimen records there appears to be a degree of habitat fidelity within the populations of all three Sceliotrachelus species. Sceliotrachelus midgleyi occurs at high altitude (1197–1805 m) across a variety of micro-habitats present in Camdebo Escarpment Thicket (Albany Thicket biome). The distribution of this species may well extend south-east and north-west where similar habitat exists, delimited by the vegetation types and altitude defining the Sneeuberg floristic centre of endemism (
Based on the sampling techniques (mostly leaf litter extraction, pitfall traps, and yellow pan traps, with only two records from Malaise traps) that were successful in collecting specimens of Afrisolia, Pulchrisolia and Sceliotrachelus, as well as their morphology, species of these genera appear to be predominantly associated with leaf litter habitat. We hypothesize that the strengthened, modified fore wings curving over the body are functionally analogous to beetle elytra (i.e. creating a protective shield), to facilitate their lifestyle in the leaf litter environment. This would be a convergent adaptation with the wing morphology present in female Acanthobetyla Dodd, 1926 (Diapriidae, Ambositrinae), which are leaf litter inhabitants. Sceliotrachelus are likely to spend most of their time on the ground and, with the exception of S. karooensis, have probably lost their ability to fly, as first suggested by
With further intense sampling, using continuous inventory surveys comprising a variety of collecting methods, we expect that more Sceliotrachelus species will be uncovered across the Cape region of South Africa. Focused biological investigation of potential host invertebrates residing in the leaf litter or soil environment is required to elucidate the biology of these enigmatic platygastrids. In terms of the knowledge of the world fauna, the majority of sceliotracheline species still remain to be discovered and formally described. Contemporary revision based on existing specimens in world collections have resulted in dramatic elevation of species richness within each genus, and given that the available specimens are representative of insufficient inventory surveys of world vegetation and habitat types, we predict that there is still a wealth of undiscovered richness at species and even generic level.
Acknowledgements
Thanks to Marlene McKay, Richard and Kitty Viljoen (Asante Sana Game Reserve); and Cape Nature reserve managers, Tom Barry (Gamkaberg Nature Reserve), and Marius Brand (Anysberg Nature Reserve) for providing permission to run long-term inventory surveys on their reserves and for logistical support in the field. Aisha Mayekiso, the late Nosiphiwo Goci, Mmamotswa Mosweu, Yvonne Samuels, the late Nkosinathi Babu, Victor Mutavhatsindi, Tiyisani Chavala, Susanna das Neves, Tiffany Wynford, and Aabid Abrahams are thanked for their dedication to processing samples at Iziko South African Museum. Dawn Larsen and Robyn Tourle are thanked for their assistance in the field during the Conservation Farming Project co-ordinated by the National Botanical Institute (now SANBI). Thanks to John Donaldson and Ingrid Nanni from SANBI for making this project a reality. Eastern Cape Nature Conservation and Cape Nature (Western Cape Province) granted collecting permits. CSIRO Publishing granted permission to reuse the Parabaeus SEM images. Thank you to the many curators of the various international insect collections for facilitating access to specimens or images of types. Scanning electron micrographs were produced by Jonathan Bremer (FDACS/DPI). Thank you to Whit Farnum, Crystal Maier, and Melissa Aja (Museum of Comparative Zoology, Harvard University) for help in obtaining images of the Sceliotrachelus braunsi holotype. Thank you to Ovidiu Popovici, Jon Bremer and Paul Skelley for their careful reviews of the manuscript. This material is based upon work supported by the South African National Research Foundation under grants GUN 61497 and GUN 98115 to S. van Noort; by the National Science Foundation under grant No. DEB–0614764 to N.F. Johnson and A.D. Austin; by GEF funding through the World Bank for the South African Conservation Farming project co-ordinated by the National Botanical Institute (now SANBI); and by the Florida Department of Agriculture and Consumer Services – Division of Plant Industry to Elijah Talamas.
References
- Alekseev VN (1979) [A new species of the genus Isolia (Hymenoptera, Platygastridae) from Central Asia.]. Zoologicheskii Zhurnal 58: 763–764. http://doi.org/10.5281/zenodo.23374
- Anjana M, Rajmohana K, Vimala D, Sundararaj R (2016) On a new species of Amitus Haldeman (Hymenoptera: Platygastridae) parasitizing whitefly Zaphanera sp. (Aleyrodidae) on Vigna trilobata from India. Halteres 7: 106–111. http://doi.org/10.5281/zenodo.55316
- Annecke DP, Prinsloo GL (1977) A new species of Allotropa Foerster (Hymenoptera: Platygasteridae) parasitic in the citrus mealybug, Planococcus citri (Risso) (Homoptera: Pseudococcidae), in South Africa. Journal of the Entomological Society of South Africa 40: 105–108.
- Anonymous (2020) S2A3 Biographical Database of Southern African Science: Brauns, Dr Johannes Heinrich Justus Carl Ernst (entomology, plant collection). http://www.s2a3.org.za/bio/Biograph_final.php?serial=343 [Accessed 6 July 2020]
- Asadi-Farfar M, Karimpour Y, Lotfalizadeh H, Caleca V (2020) New records of Sceliotrachelinae (Hymenoptera: Platygastridae) in Iran. Journal of Insect Biodiversity and Systematics 6: 125–133. http://jibs.modares.ac.ir/article-36-39536-en.html
- Ashmead WH (1893) A monograph of the North American Proctotrypidae. Bulletin of the Entomological Society of America 45: 1–472. https://doi.org/10.5479/si.03629236.45.1
- Ashmead WH (1894) A new genus and species of Proctotrypidae and a new species of Brachysticha, bred by Prof. F.M. Webster. The Journal of the Cincinnati Society of Natural History 17: 170–172.
- Ashmead WH (1901) Three new parasitic Hymenoptera from South Africa. The Canadian Entomologist 33: 138–139. https://doi.org/10.4039/Ent33138-5
- Ashmead WH (1901) Hymenoptera Parasitica. In: Sharp D (Ed.) Fauna Hawaiiensis. The University Press, Cambridge, 1: 277–364. https://doi.org/10.5962/bhl.title.4628
- Ashmead WH (1903) Classification of the pointed-tailed wasps, or the superfamily Proctotrypoidea. II. Journal of the New York Entomological Society 11: 28–35. https://doi.org/10.4039/Ent35303-11
- Austin AD (1990) A revision of Old World Parabaeus Kieffer (Hymenoptera: Platygastridae), a group of highly modified apterous parasitoids inhabiting leaf litter. Invertebrate Taxonomy 3: 647–659. https://doi.org/10.1071/IT9890647
- Austin AD, Field SA (1997) The ovipositor system of scelionid and platygastrid wasps (Hymenoptera, Platygastroidea), comparative morphology and phylogenetic implications. Invertebrate Taxonomy 11: 1–87. https://doi.org/10.1071/IT95048
- Asadi-Farfar M, Karimpour Y, Lotfalizadeh H, Caleca V (2020) New records of Sceliotrachelinae (Hymenoptera: Platygastridae) in Iran. Journal of Insect Biodiversity and Systematics 6: 125–133. http://zoobank.org/References/F5D5B569-0C77-4387-B221-416012F378F0
- Bedford ECG, Thomas ED (1965) Biological control of the citrus blackfly, Aleurocanthus woglumi (Ashby) (Homoptera: Aleyrodidae) in South Africa. Journal of the Entomological Society of Southern Africa 28: 117–132.
- Biodiversity Explorer (2020) Biodiversity explorers: Brauns, Hans Heinrich Justus Carl Ernst. http://www.biodiversityexplorer.info/people/brauns-hhjce.htm [Accessed 30 June 2020]
- Brèthes J (1914) Les ennemis de la “Diaspis pentagona” dans la Republique Argentine. Nunquam Otiosus 1914: 1–16.
- Brèthes J (1916) Hyménoptères parasites de l’Amérique méridionale. Anales del Museo Nacional de Historia Natural de Buenos Aires 27: 401–430.
- Brues CT (1908) Hymenoptera. Fam. Scelionidae. Genera Insectorum 80: 1–59.
- Brues CT (1909) Notes and descriptions of North American parasitic Hymenoptera. VII. Bulletin of the Wisconsin Natural History Society 7: 154–163.
- Brues CT (1940) Fossil parasitic Hymenoptera of the family Scelionidae from Baltic Amber. Proceedings of the American Academy of Arts and Sciences 74: 69–90. https://doi.org/10.2307/20023360
- Buhl PN (1999) A synopsis of the Platygastridae of Fennoscandia and Denmark (Hymenoptera, Platygastroidea). Entomofauna, Zeitschrift für Entomologie 20: 17–52.
- Buhl PN (2002) Contributions to the platygastrid fauna of Panama (Hymenoptera, Platygastridae). Entomofauna, Zeitschrift für Entomologie 23: 309–330.
- Buhl PN (2008) On twenty-six new or little known species of Platygastridae (Hymenoptera, Platygastroidea). Entomofauna, Zeitschrift für Entomologie 29: 385–416.
- Buhl PN (2011) Species of Platygastrinae and Sceliotrachelinae from rainforest canopies in Tanzania, with keys to the Afrotropical species of Amblyaspis, Inostemma, Leptacis, Platygaster and Synopeas (Hymenoptera, Platygastridae). Tijdschrift voor Entomologie 154: 75–126. https://doi.org/10.1163/22119434-900000309
- Buhl PN (2019) A new species of Allotropa Förster (Hymenoptera: Platygastridae), parasitoid of Delottococcus aberiae (De Lotto) (Hemiptera: Pseudococcidae) in South Africa. Entomologist’s Monthly Magazine 155: 59–64. https://doi.org/10.31184/M00138908.1551.3957
- Buhl PN, Choi JY (2006) Taxonomie Review of the family Platygastridae (Hymenoptera: Platygastroidea) from the Korean Peninsula. Journal of Asia-Pacific Entomology 9(2): 121–137. https://doi.org/10.1016/S1226-8615(08)60283-6
- Buhl PN, Notton DG (2009) A revised catalogue of the Platygastridae of the British Isles (Hymenoptera: Platygastroidea). Journal of Natural History 43: 1651–1703. https://doi.org/10.1080/00222930902993732
- Carpenter FM (1992) Arthropoda 4. Superclass Hexapoda. Treatise on Invertebrate Paleontology. Part R. Boulder, CO Geological Society of America, [i–ii,] 279–655.
- Chen H, Lahey Z, Talamas E, Valerio A, Masner L, Musetti L, Klompen H, Polaszek A, Austin A, Johnson N (2021) An integrated phylogenetic reassessment of the parasitoid superfamily Platygastroidea (Hymenoptera: Proctotrupomorpha) results in a revised familial classification. Systematic Entomology 46: 1088–1113. https://doi.org/10.1111/syen.12511
- Clark VR, Barker N, Mucina L (2009) The Sneeuberg: A new centre of floristic endemism on the Great Escarpment, South Africa. South African Journal of Botany 75: 196–238. https://doi.org/10.1016/j.sajb.2008.10.010
- Crawford JC (1916) Some new American Hymenoptera. Insecutor Inscitiae Menstruus 4: 135–144.
- Cresson ET (1887) Synopsis of the Families and Genera of the Hymenoptera of America, North of Mexico: Together with a Catalogue of the Described Species, and Bibliography. PC Stockhausen, printer. https://doi.org/10.5962/bhl.title.38716
- Crosskey RW [Ed.] (1980) Catalogue of the Diptera of the Afrotropical Region. London, 1437 pp.
- Crosskey RW, White GB (1977) The Afrotropical Region. Journal of Natural History 11: 541–544. https://doi.org/10.1080/00222937700770461
- Darlington PJ (1957) Zoogeography: the geographical distribution of animals. Wiley, New York, 675 pp.
- Dalla-Torre CG de (1898) Catalogus Hymenopterorum hucusque descriptorum systematicus et synonymicus. Volumen V: Chalcididae et Proctotrupidae. Guilelmi Engelmann, Lipsiae. [Leipzig.]: 1–598.
- Darlington PJ (1957) Zoogeography: the geographical distribution of animals. Wiley, New York, 675 pp.
- Debauche HR (1947) Scelionidae de la faune belge (Hymenoptera Parasitica). Bulletin et Annales de la Societe Entomologique de Belge 83: 255–285.
- De Santis L (1941) Lista de himenopteros parasitos y predatores de los insectos de la Republica Argentina. Boletim da Sociedade Brasileira de Agronomia 4: 1–66.
- De Santis L (1967) Catálogo de los Himenópteros Argentinos de la Serie Parasitica, incluyendo Bethyloidea. Comision de Investigacion Cientifica, La Plata, 337 pp.
- De Santis L (1970) El genero Parabaeus de Himenopteros Proctotrupoideos. Revista de la Sociedad Entomológica Argentina 32: 47–50.
- De Santis L (1971) Nuevas citas de Himenopteros para la Republica Argentina. Revista de la Facultad de Agronomía de La Plata 47: 201–207.
- De Santis L (1980) Catalogo de los Himenopteros Brasilenos de la serie Parasitica incluyendo Bethyloidea. Editora da Universidade Federal do Parana, Curitiba, 395 pp.
- Ditsong Museums of South Africa (2018) General entomology collection. http://www.ditsong.org.za/en/?exhibition=general-entomology-collection [Accessed 31 July 2018]
- Dodd AP (1913) Some new parasitic Hymenoptera from Australia. Archiv für Naturgeschichte 79: 164–182.
- Engel MS, Huang DY, Alqarni AS, Cai C, Alvarado M, Breitkreuz LCV, Azar D (2016) An apterous scelionid wasp in mid-Cretaceous Burmese amber (Hymenoptera: Scelionidae). Comptes Rendus Palevol 16: 5–11. https://doi.org/10.1016/j.crpv.2016.03.005
- Evans GA, Peña JE (2005) A new Fidiobia species (Hymenoptera: Platygastridae) reared from eggs of Diaprepes doublierii (Coleoptera: Curculionidae) from Dominica. Florida Entomologist 88: 61–66. https://doi.org/10.1653/0015-4040(2005)088[0061:ANFSHP]2.0.CO;2
- Fabritius K, Grellmann D (1971) Contributii la studiul inostemminelor din România (Hymenoptera, Platygasteridae, Inostamminae). Lucrari Stiintifice – Zoologie, Constanta 1971: 5–14.
- Fabritius K (1974) Sceliotrachelinae Brues, 1908 (Hymenoptera, Platygasteridae) neu fur die Fauna Rumaniens. Travaux du Museum d’Histoire Naturelle “Grigore Antipa” 15: 293–295.
- Forbes SA (1885) Fourteenth report of the Illinois State Entomologist. Contributions to Agricultural Entomology. Transactions of the Illinois State Department of Agriculture 22: 9–74.
- Förster A (1856) Hymenopterologische Studien. Heft II. Chalcidiae und Proctotrupii. Aachen, 1856: 1–152.
- Förster A (1878) Kleine Monographien parasitischer Hymenopleren. Verh. nat. Ver. Pr. Rheinl. Westf. 35: 42–47.
- Fouts RM (1924) Revision of the North American wasps of the subfamily Platygasterinae. Proceedings of the United States National Museum 63: 1–145. https://doi.org/10.5479/si.00963801.63-2484.1
- Ghahari H, Buhl PN (2011) Check-list of Iranian Platygastridae (Hymenoptera, Platygastroidea). Entomofauna 32: 329–336.
- Greathead DJ (1971) A review of biological control in the Ethiopian region. Commonwealth Institute of Biological Control Technical Communication No 5: 1–162.
- Haldeman SS (1850) On four new species of Hemiptera of the genera Ploiaria, Chermes and Aleurodes, and two new Hymenoptera parasitic in the last named genus. American Journal of Science 9: 108–111.
- Harris RA (1979) A glossary of surface sculpturing. California Department of Food and Agriculture, Bureau of Entomology 28: 1–31.
- He J-H, Chen X-X, Fan J-J, Li Q, Liu C-M, Lou X-M, Ma Y, Wang S-F, Wu Y-R, Xu Z-H, Xu Z-F, Yao J (2004) Hymenopteran Insect Fauna of Zhejiang. Science Press, Beijing, 1373 pp.
- Hellén W (1968) Platygasterinenfunde aus Finnland (Hymenoptera, Proctotrupoidea). Notulae Entomologicae 48: 45–48.
- Hoelmer KA, Grace JK (1989) Chapter 10 – Citrus Blackfly. In: Dahlsten DL, Garcia R (Eds) Eradication of exotic pests: analysis with case histories. Yale University Press, 147–165. https://doi.org/10.2307/j.ctt2250vh8
- Howard LO (1886) A generic synopsis of the hymenopterous family Proctotrupidae. Transactions of the American Entomological Society 13: 169–178. https://doi.org/10.2307/25076475
- Huldén L (1986) The whiteflies (Homoptera, Aleyrodoidea) and their parasites in Finland. Notulae entomologicae 66: 1–40.
- Hustache A (1929) Curculionidae. Voyage de Ch. Alluaud et R. Jeannel en Afrique Orientale (1911–1912). Résultats scentifiques, Coleóptera 30: 365–562. [+ pl.X–XI. [l-VIII-1929]]
- Jansson A (1939) Studier oever svenska proctotrupider. I. Foer faunan nya slaekten. Entomologisk Tidskrift 60: 155–175.
- Jansson A (1956) Studier over svenska proctotrupider. 10. Fidiobia tatrae Szel. (Platygasteridae, Inostemmatinae, adhuc falso Platygasterinae), en art med utbredd foerekomst i Sverige. Entomologisk Tidskrift 77: 87–90. http://doi.org/10.5281/zenodo.24441
- Kieffer JJ (1910) Diagnoses de nouveaux genres et de nouvelles espèces de Scélionides (Hym.) des Iles Séchelles. Bulletin de la Société entomologique de France 1910: 292–294. https://doi.org/10.5962/bhl.part.4689
- Kieffer JJ (1912) Hymenoptera, Proctotrupoidea. Transactions of the Linnean Society of London, 2nd series 15: 45–80. https://doi.org/10.1111/j.1096-3642.1912.tb00089.x
- Kieffer JJ (1914) Description de deux genres nouveaux et de quatre especes nouvelles de Proctotrypides (Hym.). Bulletin De La Société Entomologique De France 1914: 180–181. http://doi.org/10.5281/zenodo.24318
- Kieffer JJ (1916) Beitrag zur Kenntnis der Platygasterinae und ihrer Lebensweise. Centralblatt Fur Bakteriologie, Parasitenkunde Und Infektionskrankheiten 46: 547–592. http://doi.org/10.5281/zenodo.24361
- Kieffer JJ (1921) Un platygasterine parasite d’un cynipide. Brotéria 19: 68–69.
- Kieffer JJ (1926) Scelionidae. Das Tierreich. Vol. 48. Walter de Gruyter & Co., Berlin, 885 pp.
- Kock WJ, Krüger DW [Eds] (1972) Brauns, Justus Carl Ernst Johannes. In: Dictionary of South African Biography, 2. Tafelberg Publishers, Cape Town, 84–86.
- Kozlov MA (1970) [Supergeneric groupings of Proctotrupoidea (Hymenoptera)]. Entomologicheskoye Obozreniye 49: 203–226. [In Russian]
- Kozlov MA (1971) Proctotrupoids (Hymenoptera, Proctotrupoidea) of the USSR. Proceedings of the All-Union Entomological Society 54: 3–67. [In Russian]
- Kozlov MA (1972) [On the fauna of Hymenoptera Proctotrupoidea of the Mongolian People’s Republic. I. Heloridae, Proctotrupidae, Scelionidae. ] Insects of Mongolia 1: 645–672. https://dx.doi.org/10.5281/zenodo.23826 [In Russian]
- Kozlov MA (1977) [A new genus and species of parasitoid of the family Platygastridae (Hymenoptera, Proctotrupoidea) from Armenia. ] Trudy Zoologicheskogo Instituta Akademii Nauk SSSR 64: 79–81. [In Russian]
- Kozlov MA (1978) [Superfamily Proctotrupoidea]. In: Medvedev GS (Ed.) [Determination of insects of the European portion of the USSR. ] Vol. 3, part 2. Nauka, Leningrad, 538–664. [In Russian]
- Kozlov MA (1995) [26. Fam. Platygastridae.] In: Lehr PA (Ed.) [Key to insects of Russian Far East in six volume. vol. 4. Neuropteroidea, Mecoptera, Hymenoptera. Part 2. Hymenoptera.] Dal’nauka, Vladivostok, 121–127. [In Russian]
- Krombein KV, Burks BD (1967) Hymenoptera of America North of Mexico: Synoptic Catalog, 2nd Supplement. US Department of Agriculture, 584 pp.
- Lahey Z, Masner L, Johnson NF (2019a) Calixomeria, a new genus of Sceliotrachelinae (Hymenoptera, Platygastridae) from Australia. ZooKeys 830: 63–73. https://doi.org/10.3897/zookeys.830.32463
- Lahey Z, van Noort S, Polaszek A, Masner L, Johnson N (2019b) Revision of the Afrotropical genus Pulchrisolia Szabó (Hymenoptera, Platygastridae, Sceliotrachelinae). In: Talamas E (Ed.) Advances in the Systematics of Platygastroidea II. Journal of Hymenoptera Research 73: 39–71. https://doi.org/10.3897/jhr.73.33876
- Lahey Z, Masner L, Johnson NF, Polaszek A (2019c) Revision of Aleyroctonus Masner & Huggert (Hymenoptera, Platygastridae, Sceliotrachelinae). In: Talamas E (Ed.) Advances in the Systematics of Platygastroidea II. Journal of Hymenoptera Research 73: 73–93. https://doi.org/10.3897/jhr.73.38383
- Lahey Z, Talamas E, Masner L, Johnson NF (2021) Revision of the Australian genus Alfredella Masner & Huggert (Hymenoptera, Platygastridae, Sceliotrachelinae). In: Lahey Z, Talamas E (Eds) Advances in the Systematics of Platygastroidea III. Journal of Hymenoptera Research 87: 81–113. https://doi.org/10.3897/jhr.87.58368
- Latreille PA (1802) Histoire naturelle, générale et particuliére des crustacés et des insectes. Vol. 13. F. Dufart, Paris, 432 pp.
- Lemarie J (1958) Contribution to knowledge of the parasites of the pine bud-moth Exoteleia (Heringia) dodecella L. Part I. Chalcidoidea, Proctotrupoidea, Bethyloidea. Zoologicke Listy 21: 221–230.
- Lemarie J (1961) Contribution to knowledge of the parasites of the pine bud-moth Exoteleia (Heringia) dodecella L. Part 4. Ichneumonidae, Braconidae, Chalcidoidea, Bethyloidea. Zoologicke Listy 24: 119–126.
- Loiácono MS, Margaría CB (2002) Systematics, morphology and physiology. Ceraphronoidea, Platygastroidea and Proctotrupoidea from Brazil (Hymenoptera). Neotropical Entomology 31: 551–560. https://doi.org/10.1590/S1519-566X2002000400007
- Lotfalizadeh H (2018) Four subfamilies of Platygastroidea (Hymenoptera) in the northwest of Iran with new report of Sceliotrachelinae. North-Western Journal of Zoology 14: 237–242.
- MacGown M, Nebeker T (1978) Taxonomic review of Amitus (Hymenoptera: Proctotrupoidea, Platygastridae) of the western hemisphere. Canadian Entomologist 110: 275–283. https://doi.org/10.4039/Ent110275-3
- Madl M (2016) A catalogue of the family Platygastridae (Platygastroidea) of the Malagasy subregion. Part I: Subfamilies Platygastrinae and Sceliotrachelinae (Insecta: Hymenoptera). Linzer Biologische Beitraege 48: 55–69.
- Maneval H (1940) Fam. XVII. Proctotrypides. In: Perrier R (Ed.) La Faune de la France en tableaux synoptiques illustrés. Tome VII. Hyménoptères par Lucien Berland avec la collaboration de MM. Raymond Benoit, Francis Bernard, Henri Maneval, Paris, 93–118.
- Mani MS (1941) Serphoidea. Catalogue of Indian Insects 26: 1–60.
- Mani MS, Sharma SK (1982) Proctotrupoidea (Hymenoptera) from India. A review. Oriental Insects 16: 135–258. https://doi.org/10.1080/00305316.1982.10434314
- Marshall TA (1873) A catalogue of British Hymenoptera; Oxyura. Entomological Society of London, London, 27 pp.
- Masner L (1958) An interesting new genus of Scelionidae from S.W. Africa (Hymenoptera: Proctotrupoidea). Proceedings of the Royal Entomological Society of London (B) 27: 101–104. https://doi.org/10.1111/j.1365-3113.1958.tb00420.x
- Masner L (1964) Remarks on Sceliotrachelus Brues and allied genera (Hymenoptera, Platygasteridae). Psyche 71: 8–11. https://doi.org/10.1155/1964/40282
- Masner L (1965) The types of Proctotrupoidea (Hymenoptera) in the British Museum (Natural History) and in the Hope Department of Entomology, Oxford. Bulletin of the British Museum (Natural History) Entomology Supplement 1: 1–154. https://doi.org/10.5962/p.97756
- Masner L (1968) A new genus of Scelionidae (Hymenoptera) with austral disjunctive distribution. New Zealand Journal of Science 11: 652–663. http://doi.org/10.5281/zenodo.23721
- Masner L (1976) Revisionary notes and keys to world genera of Scelionidae (Hymenoptera: Proctotrupoidea). Memoirs of the Entomological Society of Canada, 108: 1–87. https://doi.org/10.4039/entm10897fv
- Masner L (1993) Superfamily Platygastroidea. In: Goulet H, Huber J (Eds) Hymenoptera of the World: an identification guide to families. Research Branch, Agriculture Canada, Ottawa, Canada, 558–565.
- Masner L, Johnson NF (2007) Janzenella, an enigmatic new genus of scelionid wasp from Costa Rica (Hymenoptera: Platygastroidea, Scelionidae). American Museum Novitates 3574: 1–7. https://doi.org/10.1206/0003-0082(2007)3574[1:JAENGO]2.0.CO;2
- Masner L, Johnson NF, Polaszek A (2007) Redescription of Archaeoscelio Brues and description of three new genera of Scelionidae (Hymenoptera): a challenge to the definition of the family. American Museum Novitates 3550: 1–24. https://doi.org/10.1206/0003-0082(2007)3550[1:ROABAD]2.0.CO;2
- Masner L, Huggert L (1989) World review and keys to genera of the subfamily Inostemmatinae with reassignment of the taxa to the Platygastrinae and Sceliotrachelinae (Hymenoptera: Platygastridae). Memoirs of the Entomological Society of Canada 147: 1–214. https://doi.org/10.4039/entm121147fv
- Masner L, Muesebeck CF (1968) The types of Proctotrupoidea (Hymenoptera) in the United States National Museum. Bulletin of the United States National Museum 270: 1–143. https://doi.org/10.5479/si.03629236.270
- McKellar RC, Engel MS (2012) Hymenoptera in Canadian Cretaceous amber (Insecta). Cretaceous Research 35: 258–279. https://doi.org/10.1016/j.cretres.2011.12.009
- Mikó I, Vilhelmsen L, Johnson NF, Masner L, Pénzes Z (2007) Skeletomusculature of Scelionidae (Hymenoptera: Platygastroidea): head and mesosoma. Zootaxa 1571: 1–78. https://doi.org/10.11646/zootaxa.1571.1.1
- Morley C (1929) Catalogus Oxyurarum Britannicorum. Transactions of the Suffolk Naturalists’ Society 1: 39–60.
- Mucina L, Rutherford MC [Eds] (2006) The vegetation of South Africa, Lesotho and Swaziland. SANBI, Pretoria, 807 pp.
- Muesebeck CFW (1979) Family Platygastridae. In: Krombein K, Hurd Jr PD, Smith DR, Burks BD (Eds) Catalog of Hymenoptera in America North of Mexico. Vol. 1. Smithsonian Institution Press, Washington DC, 1171–1186.
- Muesebeck CFW, Masner L (1967) Proctotrupoidea. In: Krombein KV, Burks BD (Eds) Hymenoptera of America north of Mexico. Synoptic catalog. Agriculture Monogr. 2 (Suppl. 2), 285–305.
- Muesebeck CFW, Walkley LM (1956) Type species of the genera and subgenera of parasitic wasps comprising the superfamily Proctotrupoidea (order Hymenoptera). Proceedings of the United States National Museum 105: 319–419. https://doi.org/10.5479/si.00963801.3359.319
- Murphy NP, Carey D, Castro LR, Dowton M, Austin AD (2007) Phylogeny of the platygastroid wasps (Hymenoptera) based on sequences from the 18S rRNA, 28S rRNA and cytochrome oxidase genes: implications for the evolution of the ovipositor system and host relationships. Biological Journal of the Linnean Society 91: 653–669. https://doi.org/10.1111/j.1095-8312.2007.00825.x
- Nguyen R (1988) Amitus hesperidum (Hymenoptera: Platygasteridae), a parasite of the citrus blackfly (Aleurocanthus woglumi). Florida Department of Agricultural Consumer Services, Division P1 Industry, Entomolgy Circular 311: 1–2.
- Nixon GEJ (1942) A remarkable new genus of Telenominae (Hym., Proctotrupoidea, Scelionidae). Annals and Magazine of Natural History 9: 462–464. https://doi.org/10.1080/03745481.1942.9755497
- Nixon GEJ (1969) Two new species of Platystasius Nixon with a note on the generic relationship between Platystasius and Fidiobia (Hymenoptera: Platygasteridae). Proceedings of the Entomological Society of Washington 71: 445–449.
- Notton DG (2010) A catalogue of the types of Platygastridae (Hymenoptera, Platygastroidea) at the Muséum national d'Histoire naturelle, Paris, with brief notes on the history of the collection. Zootaxa 2358: 1–24. https://doi.org/10.11646/zootaxa.2358.1.1
- Notton DG, Popovici OA, van Achterberg C, de Rond J, Burn JT (2014) Parasitoid wasps new to Britain (Hymenoptera: Platygastridae, Eurytomidae, Braconidae & Bethylidae). European Journal of Taxonomy 99: 1–20. https://doi.org/10.5852/ejt.2014.99
- Oglobin AA (1944) Two new species of Proctotrupoidea from Iowa (Hymenoptera). Proceedings of the Entomological Society of Washington 46: 155–158.
- Ortega-Blanco J, McKellar RC, Engel MS (2014) Diverse scelionid wasps in Early Cretaceous amber from Spain (Hymenoptera: Platygastroidea). Bulletin of Geosciences 89: 553–571. https://doi.org/10.3140/bull.geosci.1463
- Polaszek A (1997) Amitus Haldeman (Hymenoptera: Platygastridae): a genus of whitefly parasitoids new to Britain. Entomologist’s Monthly Magazine 133: 77–79.
- Polaszek A (2009) Masnerium Polaszek 2009, gen. n. Masnerium wellsae gen. n., sp. n. (hymenoptera, Platygastridae, Sceliotrachelinae) a parasitoid of Aleuroduplidens wellsae Martin (Hemiptera, Aleyrodidae) in Australia. Zookeys 20: 119–125. https://doi.org/10.3897/zookeys.20.189
- Popovici O, Buhl PN (2010) The West Palearctic species of Fidiobia Ashmead, 1894 (Hymenoptera: Platygastroidea). Journal of Natural History 44: 1131–1164. https://doi.org/10.1080/00222931003632740
- Popovici OA, Vilhelmsen L, Masner L, Mikó I, Johnson N (2017) Maxillolabial complex in scelionids (Hymenoptera: Platygastroidea): morphology and phylogenetic implications. Insect Systematics & Evolution 48: 1–125. https://doi.org/10.1163/1876312X-48022156
- Rajmohana K (2006) Studies on Proctotrupoidea and Platygastroidea (Hymenoptera: Insecta) of Kerala. Memoirs of the Zoological Survey of India 21: 1–153. https://doi.org/10.11609/JoTT.ZPJ.1570.2506-13
- Rajmohana K (2014) A systematic inventory of Scelioninae and Teleasinae (Hymenoptera: Platygastridae) in the rice ecosystems of north-central Kerala. Memoirs of Zoological Survey of India 22: 1–72.
- Risbec J (1955a) Les parasites de Pseudococcus njalensis Laing et de Pseudococcus bingervillensis Magnin. L’Agronomie Tropicale 10: 231–237.
- Risbec J (1955b) Platygasterinae de Madagascar (Hym. Proctotrupidae). Bulletin de la Société Zoologique de France 80: 109–118.
- Risbec J (1957) Description d’un nouveau Baeinien africain Parabaeus machadoi n. sp. (Hym. – Scelionidae). Publicacoes Culturais da Companhia de Diamantes de Angola 34: 87–89.
- Rutherford MC, Mucina L, Powrie LW (2006) Biomes and bioregions of Southern Africa. In: Mucina L, Rutherford MC (Eds) The vegetation of South Africa, Lesotho and Swaziland. SANBI, Pretoria, 30–51.
- Sarazin MJ (1986) Primary types of Ceraphronoidea, Evaniodea, Proctotrupoidea, and Trigonaloidea (Hymenoptera) in the Canadian National Collection. The Canadian Entomologist 118: 957–989. https://doi.org/10.4039/Ent118957-10
- Sclater PL (1858) On the general geographical distribution of the members of the class Aves. Journal of the Proceedings of the Linnean Society, Zoology 2: 130–145. https://doi.org/10.1111/j.1096-3642.1858.tb02549.x
- Sharkey MJ (2007) Phylogeny and classification of Hymenoptera. In: Zhang Z-Q, Shear WA (Eds) Linnaeus tercentenary: progress in invertebrate taxonomy. Zootaxa 1668: 521–548. https://doi.org/10.11646/zootaxa.1668.1.25
- Silvestri F (1927) Contribuzione alla conoscenza degli Aleurodidae (Insecta: Hemiptera) viventi su citrus in estremo oriente e dei loro parassiti (Hym.). Bollettino del Laboratorio di zoologia generale e agraria della R. Scuola superiore d’agricoltura in Portici 21: 1–60.
- Szabó JB (1958) Erganzende Beobachtungen uber die holarktische Gattung Fidiobia Ashmead 1894. Folia Entomologica Hungarica 11: 457–464.
- Szabó JB (1959) Notes on the new tribus Amitini with the descriptions of a new genus and some new species of the Arctogaea (Hymenoptera, Proctotrupoidea. Platygasteridae). Annales Musei historico-naturalis hungarici 51: 389–396.
- Szabó JB (1962) Untersuchungen an palaearktischen Proctotrupiden. I–IV. (Hymenoptera). Folia Entomologica Hungarica 15: 221–243.
- Szelényi G (1938) Über eine verkannte Gattung der Platygasteriden, nebst Beschreibung 2 neuer Arten aus der Familie der Scelioniden (Hym. Proctotr.). Folia Entomologica Hungarica 3: 100–106.
- Talamas E, Buffington M (2015) Fossil Platygastroidea in the National Museum of Natural History, Smithsonian Institution. Journal of Hymenoptera Research 47: 1–52. https://doi.org/10.3897/JHR.47.5730
- Talamas E, Johnson NF, Shih C, Ren D (2019) Proterosceliopsidae: A New Family of Platygastroidea from Cretaceous Amber. In: Talamas E (Ed.) Advances in the Systematics of Platygastroidea II. Journal of Hymenoptera Research 73: 3–38. https://doi.org/10.3897/jhr.73.32256
- Tomsík B (1950) Ceskoslovensti Inostemmini – cizopasnici u bejlomorek. Czeckoslovak Inostemmini – the parasites of Cecidomyidae. Entomologicke Listy 13: 49–64.
- van Noort S, Buffington ML, Forshage M (2015) Afrotropical Cynipoidea (Hymenoptera). ZooKeys 493: 1–176. https://doi.org/10.3897/zookeys.493.6353
- Veenakumari K, Popovici OA, Buhl PN, Mohanraj P (2018) Revision of Indian Species of Fidiobia Ashmead (Platygastridae: Sceliotrachelinae). Annales Zoologici 68: 553–600. https://doi.org/10.3161/00034541ANZ2018.68.3.013
- Veenakumari K, Rajmohana K, Manickavasagam S, Mohanraj P (2011) On a new genus of Teleasinae (Hymenoptera: Platygastridae) from India. Biosystematica 5: 39–46.
- Veenakumari K, Buhl PN, Mohanraj P (2019) Review of the genus Isolia Förster (Platygastroidea: Platygastridae: Sceliotrachelinae) with description of two new species from India. Zootaxa 4565: 451–474. https://doi.org/10.11646/zootaxa.4565.4.1
- Viggiani G, Mazzone P (1982) The Amitus Hald. (Hym. Platygastridae) of Italy, with descriptions of three new species. Journal Bollettino del Laboratorio di Entomologia Agraria “Filippo Silvestri” Portici 39: 59–69.
- Vlug HJ (1995) Catalogue of the Platygastridae (Platygastroidea) of the world (Insecta: Hymenoptera). Hymenopterorum Catalogus 19: 1–168. https://zenodo.org/record/24358
- Wallace AR (1876) The geographical distribution of animals: with a study of the relations of living and extinct faunas as elucidating the past changes of the earth’s surface. Macmillan, London, 503 pp. https://doi.org/10.5962/bhl.title.46581
- Wheatley PE (1964) The successful establishment of Eretmocerus serius Silv. (Hymenoptera: Eulophidae) in Kenya. East African Agricultural Forestry Journal 29: 236. https://doi.org/10.1080/00128325.1964.11661929