Research Article |
|
Corresponding author: Ilgoo Kang ( ikang1@lsu.edu ) Academic editor: Jose Fernandez-Triana
© 2022 Ilgoo Kang, James B. Whitfield, Brittany E. Owens, Junyan Chen.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Kang I, Whitfield JB, Owens BE, Chen J (2022) Resurrection of Neocardiochiles Szépligeti, 1908 (Hymenoptera, Braconidae, Cardiochilinae) with descriptions of five new species from the Neotropical region. Journal of Hymenoptera Research 91: 41-68. https://doi.org/10.3897/jhr.91.84937
|
Abstract
Neocardiochiles Szépligeti, 1908, is a rare Neotropical genus of the subfamily Cardiochilinae Ashmead, 1900. The genus was previously synonymized with Heteropteron Brullé, 1846 by
Keywords
Delta, DNA, Intkey, parasitoid wasps, phylogeny, taxonomy, 16S rRNA, 28S rRNA
Introduction
Members of the cardiochiline genera, Heteropteron Brullé, 1846, Neocardiochiles Szépligeti, 1908 and Wesmaelella Spinola, 1851 are rarely-collected braconid wasps in the Neotropical region, which possess a relatively large body size (6.5–13 mm). These three genera have been treated as the most plesiotypic members of the Cardiochilinae Ashmead, 1900 based on morphological characters (
Despite the rarity of the specimens of the three genera in museum collections, their relationships have been frequently discussed and have fluctuated even in recent decades (
Materials and methods
Specimen information
The type specimens of recorded species for this work were borrowed from the Hungarian Natural History Museum (
Morphological analyses
A Leica MZ75 stereomicroscope was used to examine specimens. The morphological terms are mostly based on
Molecular analyses
DNA was extracted from one to two legs of each specimen using the DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany). Mitochondrial 16S rRNA (16S), and nuclear 28S rRNA (28S) genes were targeted and amplified using the primers listed in Table
| Target genes | Primer sequences (5’→3’) | References |
|---|---|---|
| 16S rRNA | cacctgtttatcaaaaacat (F) |
|
| cttaattcaacatcgaggtc (R) |
|
|
| 28S rRNA | agagagagttcaagagtacgtg (F) |
|
| ttggtccgtgtttcaagacgg (R) |
|
|
| tagttcaccatctttcgggtc (R) |
|
Phylogenetic analyses
Morphological data
Thirty-nine morphological characters for ten Neocardiochiles species were selected and included in the phylogenetic analysis (Table

Maximum Parsimony (MP) phylogeny based on morphological data indicating the relationships of Heteropteron, Neocardiochiles and Wesmaelella. Protomicroplitis calliptera (Hymenoptera: Braconidae: Microgastrinae) is included as the outgroup. Synapomorphies are mapped on the phylogeny. Black bars indicate non-homoplastic characters, and white bars represent homoplastic characters. Characters are listed above bars, and character states are indicated below bars.
| Number | Characters | Character states |
|---|---|---|
| 1 | Y-shaped suture on frons | absent = 1; present = 2 |
| 2 | POD | broad = 1; narrow = 2 |
| 3 | Malar space width | as long as basal width of mandible = 1; shorter than basal width of mandible = 2 |
| 4* | Mouthparts length | short = 1; elongate = 2 |
| 5 | 3rd maxillary palpi shape | moderately swollen apically in lateral view = 1; strongly swollen apically in lateral view = 2 |
| 6 | Scutellar sulcus shape | transversely straight = 1; transversely curved = 2 |
| 7 | Scutellar sulcus depth | shallow and evenly impressed = 1; medially shallow, laterally deep = 2 |
| 8 | Scutellum length | longer than basal width of scutellum = 1; shorter than basal width of scutellum = 2 |
| 9 | Metascutellum length | long = 1; short = 2 |
| 10 | Posterior margin of axilla | meeting lateral margin of scutellum with narrow angle = 1; meeting lateral margin of scutellum with broad angle = 2 |
| 11 | Pronotal carina | absent = 1; present = 2 |
| 12 | Episternal scrobe | apparent = 1; weakly impressed = 2 |
| 13 | Ventral margin of metapleuron | without carinate margin = 1; anteriorly carinate = 2; entirely carinate = 3 |
| 14* | Median longitudinal furrow on propodeum | absent = 1; present = 2 |
| 15 | Curved submedial carina on propodeum | absent = 1; present posteriorly = 2 |
| 16 | Lateral margin of propodeum | absent = 1; carinate = 2 |
| 17* | Fore wing 1r | present = 1; absent = 2 |
| 18 | Fore wing 2RS shape | angled = 1; basally weakly curved = 2; straight = 3 |
| 19* | Fore wing 3RSb | broken basally = 1; evenly present = 2 |
| 20 | Fore wing (RS+M)b length | shorter than m-cu = 1; longer than m-cu = 2 |
| 21 | Fore wing (RS+M)b angle | meeting 2M with ~140° = 1; meeting 2M with 180° = 2 |
| 22 | Fore wing RS2 | present as basal stump = 1; absent = 2 |
| 23 | Fore wing r-m length | 0.5 × longer than height of second submarginal cell = 1; 0.3 × longer than height of second submarginal cell = 2; as long as height of second submarginal cell = 3 |
| 24 | Fore wing 1cu-a origin | arising from middle of 1CU = 1; arising from basal fourth of 1CU = 2 |
| 25* | Fore wing 1a | absent = 1; present = 2 |
| 26 | Hind wing M+CU length | slightly longer than 1M = 1; shorter than 1M = 2; as long as 1M = 3 |
| 27 | Hind wing cu-a length | as long as 1M = 1; shorter than 1M = 2 |
| 28* | Hind wing 2-1A length | reaching at basal half = 1; not reaching at basal half = 2 |
| 29 | Second tarsomere of fore leg length | shorter than fifth tarsomere = 1; as long as fifth tarsomere = 2 |
| 30 | Second tarsomere of middle leg length | shorter than combined length of third and fourth tarsomeres = 1; longer than combined length of third and fourth tarsomeres = 2 |
| 31 | Basal spur on hind tibia length | ~0.33 × longer than hind basitarsus = 1; ≥ 0.40 × longer than hind basitarsus = 2 |
| 32* | Tarsal claws | Simple = 1; pectinate = 2 |
| 33* | T1 ratio | ≤ 1.70 × = 1; ≥ 2.0 × = 2 |
| 34 | First laterotergite | weakly curved posteriorly = 1 strongly curved posteriorly = 2 |
| 35 | Spiracle of first laterotergite | touching dorsal margin of first laterotergite = 1; located near median = 2 |
| 36 | Spiracle of second laterotergite | close to anterior margin, but not touching = 1; located near median = 2 |
| 37* | Ovipositor sheath length | longer than hind femur = 1; shorter than hind femur; unknown = ? |
| 38 | Ovipositor sheath shape | ventro-apically round = 1; ventro-apically pointed = 2; unknown=? |
| 39* | Median longitudinal fold on hypopygium | absent = 1; present = 2; unknown = ? |
Molecular data
Maximum likelihood analysis (ML) was conducted using MEGA11 (
Results and discussion
Generic relationships and character discussion (Fig. 1 )
The Maximum Parsimony (MP) phylogenetic analysis based on morphological characters was conducted due to the issues with Heteropteron and Wesmaelella specimens mentioned above, and the limited molecular data obtained from only a few specimens of Neocardiochiles. In the MP consensus tree, Protomicroplitis was set as outgroup, and all the three ingroup genera are recovered as monophyletic groups. Based on the results, we resurrect Neocardiochiles to the generic level. A clade including Wesmaelella represented by Wesmaelella nigripennis (Szépligeti, 1902) was supported by eight synapomorphies (10-1; 13-2; 17-1; 20-1; 22-1; 33-1; 35-1; 36-1) and one homoplastic character (24-1) and recovered as the most plesiotypic member. 1r on fore wing (17-1) and RS2 (22-1) are easily observable characters to distinguish Wesmaelella from the members of Heteropteron and Neocardiochiles. A clade containing Heteropteron and Neocardiochiles were supported by six synapomorphies (1-2; 2-2; 21-2; 23-2; 25-2; 26-2) and five homoplastic characters (6-2 7-2 12-2 18-2 27-2). Three synapomorphies (13-1; 29-2; 30-2) and two homoplastic characters (19-2; 37-1) supported a clade with Heteropteron represented by the undescribed species. All the three unambiguous synapomorphies along with the ovipositor character (37-1; note: the undescribed species of Heteropteron possess a distinctively longer and more sinuate ovipositor than the members of Wesmaelella and Neocardiochiles) are easily visible diagnostic characters to identify Heteropteron. A clade including nine species of Neocardiochiles was supported by six synapomorphies (5-2; 8-2; 14-2; 15-2; 32-2; 34-2) and seven homoplastic characters (3-2; 9-2; 11-2; 16-2; 18-3; 28-2; 31-2) (Fig.
Molecular data
We did not attempt to obtain molecular data from Heteropteron and Wesmaelella specimens because they were collected in the early 1900s and/or were type specimens. Neocardiochiles specimens collected from late 1990s to early 2010s were used to obtain molecular data. Among nine species of Neocardiochiles, 16S sequences of two species, N. chriscarltoni sp. nov. and N. victoriae sp. nov., and 28S sequences of three species, N. alexeyi sp. nov., N. hasegawai comb. nov., and N. victoriae sp. nov., were obtained. Unfortunately, attempts to obtain DNA sequences from the other five species of Neocardiochiles failed. In the genetic distance analyses and maximum likelihood analyses, ~465 bp of 16S sequences and ~420 bp of 28S sequences were utilized as the final dataset, respectively. The length of 28S sequences used in the analysis was shorter than our target length because only the forward strand of N. hasegawai was successfully obtained. Interspecific genetic distance between N. chriscarltoni sp. nov. and N. victoriae sp. nov. was 12.1% for 16S (Table
| N. chriscarltoni sp. nov. | N. victoriae sp. nov. | P. calliptera (outgroup) | |
|---|---|---|---|
| N. chriscarltoni sp. nov. | 0 | ||
| N. victoriae sp. nov. | 0.121 | ||
| P. calliptera (outgroup) | 0.101 | 0.116 | 0 |
Taxonomy
Neocardiochiles
Heteropteron
Brullé, 1846, synonymized by Dangerfield, Austin, and Whitfield (1999) and confirmed by
Heteropteron whitfieldi Mercado, 2003 to Neocardiochiles whitfieldi (Mercado, 2003); Heteropteron kidonoi Dabek & Whitfield, 2020 to Neocardiochiles kidonoi (Dabek & Whitfield, 2020); Heteropteron hasegawai Dabek & Whitfield, 2020 to Neocardiochiles hasegawai (Dabek & Whitfield, 2020). New combinations.
Type species
Neocardiochiles fasciipennis Szépligeti, 1908.
Diagnosis
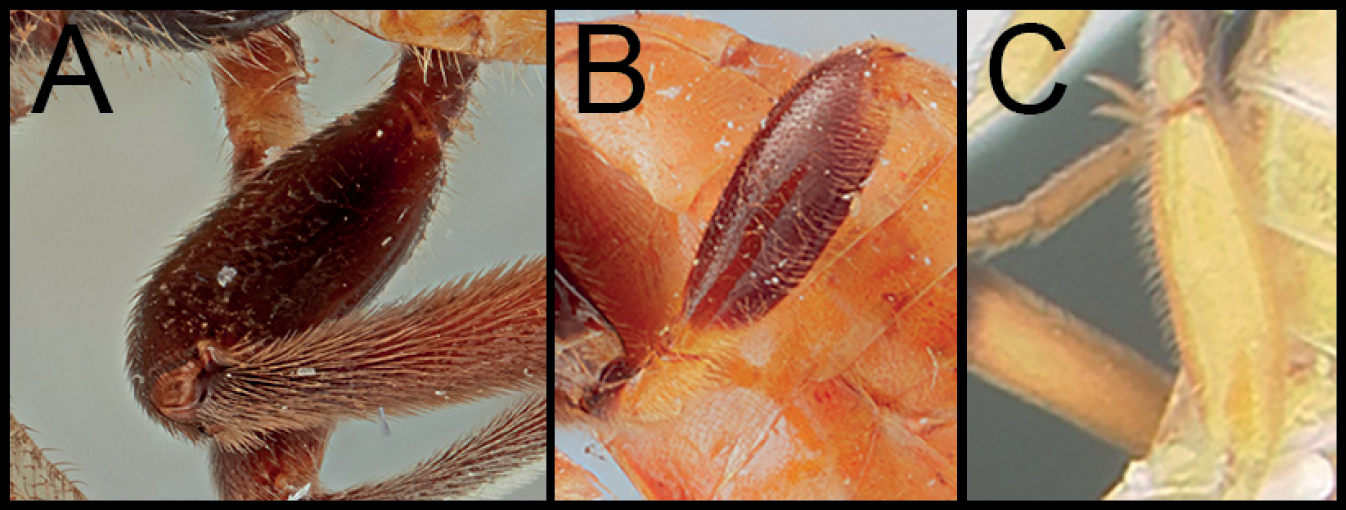
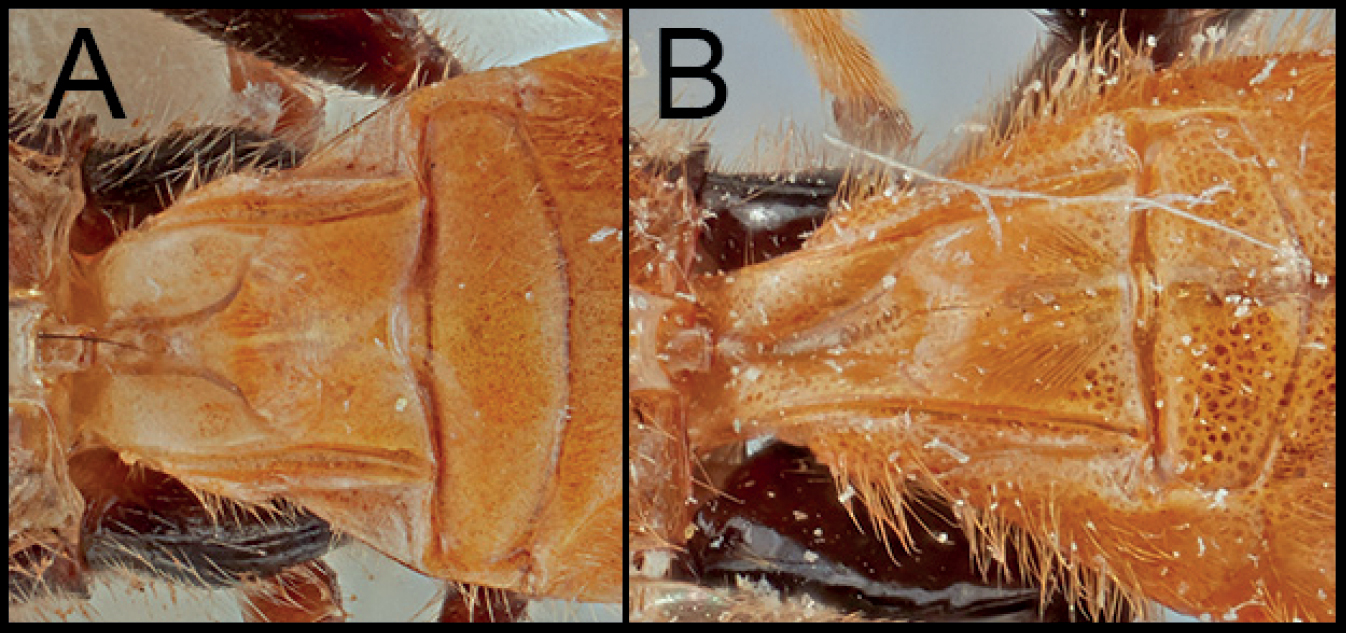
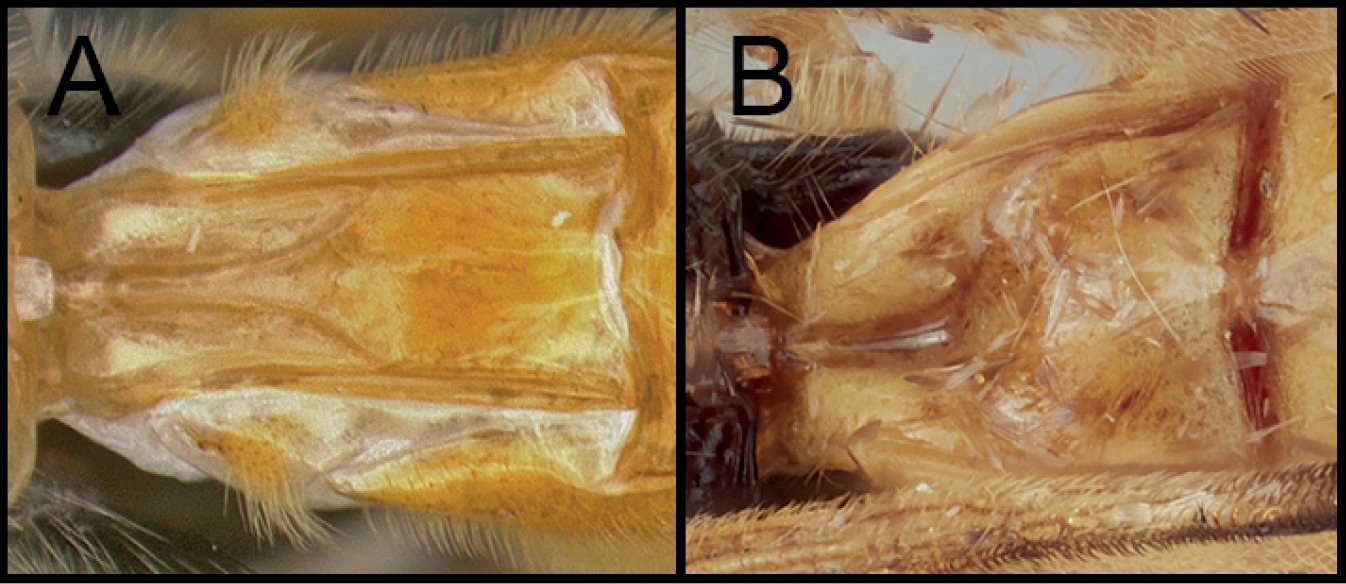
Neocardiochiles is most similar to the genera Heteropteron and Wesmaelella and shares the following characters: eyes without setae; median areola on propodeum absent; notauli weekly impressed and posteriorly absent; scutellar sulcus without any crenula. However, members of Neocardiochiles differ from Heteropteron and Wesmaelella by possessing pectinate claws; propodeum with median longitudinal furrow (Figs
Description
Body 6.5–11 mm. Head: Antenna 34–40-segmented. Face width 1.36–1.73 × longer than its height. Interantennal space with well-developed median carina. Width of anterior ocellus 0.96–1.15 × longer than POL. Eyes bulged and without interommatidial setae (Figs
Distribution
Neotropical region: Costa Rica, Ecuador, French Guiana, Mexico, and Suriname.
Biology
The two species for which hosts are known attack pyralid and depressariid caterpillars on Roupala (Proteaceae) (
Diversity
Nine species (
Key to species of Neocardiochiles of the New World
| 1 | A. Hind femur entirely melanic | 2 |
| – | B. Hind femur bicolored | 4 |
| – | C. Hind femur entirely pale | 6 |
|
||
| 2(1) | A. Fore wing entirely infuscate | N. whitfieldi |
| – | B. Fore wing two-banded | 3 |

|
||
| 3(2) | A. T1 ~ 1.06 × longer than its posterior width | N. chriscarltoni sp. nov. |
| – | B. T1 ~ 1.63 × longer than its posterior width | N. braeti sp. nov. |
|
||
| 4(1) | A. Mesoscutum entirely dark | N. fasciipennis |
| – | B. Mesoscutum entirely pale | 5 |
|
||
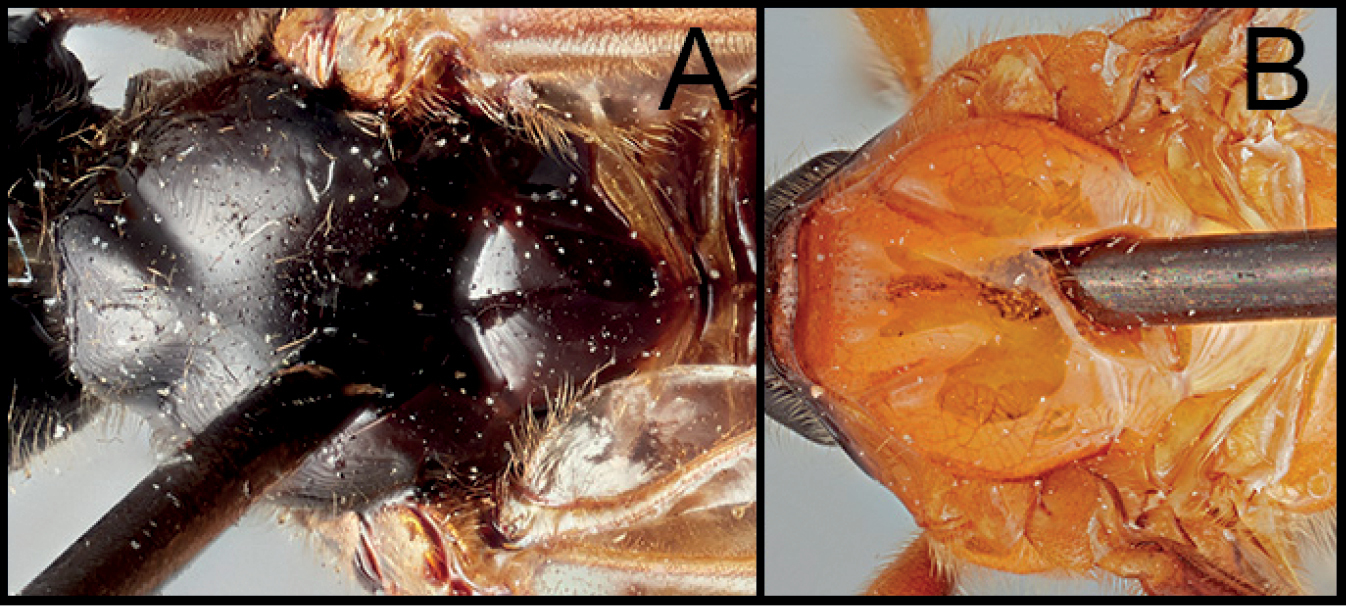
| 5(4) | A. Hind tibia mostly pale | N. kidonoi |
| – | B. Hind tibia mostly dark | N. victoriae sp. nov. |

|
||
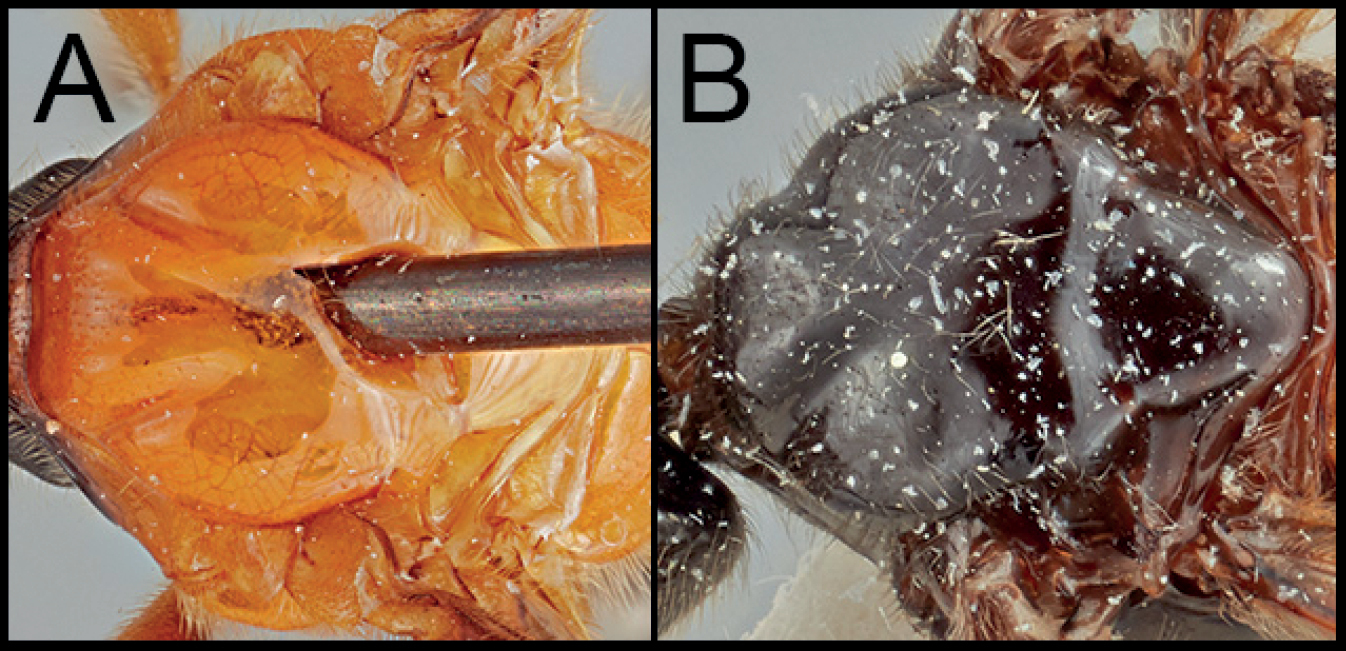
| 6(1) | A. Mesoscutum entirely pale | N. hasegawai |
| – | B. Mesoscutum entirely dark | 7 |
|
||
| 7(6) | A. T1 ~ 2.22 × longer than its posterior width, nearly rectangle | N. alexeyi sp. nov. |
| – | B. T1 ~ 1.09 × longer than its posterior width, trapezoid | N. franki sp. nov. |
|
||
Interactive key
To use the interactive key to species of Neocardiochiles in the New World (Suppl. material
- Download the zipfile of the ALA version of DELTA Editor (Open DELTA 1.02 beta) (
Dallwitz et al. 1999 ). Extract into the ‘C’ drive. The extracted folder name will be ‘open-delta-1.02-bin’. Java is required to operate DELTA Editor and Intkey. - Download the folder (‘Neocardiochiles_DELTA_Key’), which includes all source files and images of the interactive key, using the following link (https://drive.google.com/drive/folders/1aLS3vzdK52uMV0702IopDWgXaSllpJQm?usp=sharing).
- Extract the zipfile of ‘Neocardiochiles_DELTA_Key’ into the ‘open-delta-1.02-bin’ folder.
- In the folder ‘Neocardiochiles_DELTA_Key’, open the file ‘Neocardiochiles_Intkey’.
- Users can then identify described species of Neocardiochiles using this interactive key.
Species descriptions
Neocardiochiles alexeyi , sp. nov.
Material examined
Holotype Ecuador • ♀; Yasuni Research Station, Yasuni National Park, Orellana province; 00°40.4'S, 76°23.861'W; 18–24.vii.2008; A. Tishechkin; AT 853MS-2; H16634. Will be deposited in
Diagnosis
Neocardiochiles alexeyi sp. nov. can be distinguished from other members of the Neocardiochiles by the combination of the following characters: antenna 34-segmented and with pale apex (Fig.

Molecular data
28S sequences (GenBank accession number: ON040823.1);
ATGAGGAGATTCACTGTTAGCATTACTAGTATTAATGCAAATTATGATATGATTATATGATCCTTGTGGTCACAATTATTATACTTATTTGTATTATTTTATTGGTTTTGTCAGCATGCACTTCTCCTCTAGTAGAACGTCGCGACCCGTTGAATGTTTATTTATGAGCCACATGGTAGTCTTATGTATTTTATACGCAAGACCAGTGAATTTCTAATAAACTATTTGACGGTATCTAAAATGGTATTGAGCCGCAAATTTTTTTTGCGTTAGATTTATTACAAGCTAGACTTACTTTAAGCAGTACGAATTTTATGTCGTCGTTTAAACTAGTCTGCTGTTAGTGATAATATCTTTAACTGGCTTAATTTTACCGGTCAGCGATGCTACTGCTTTGGGTACTTACAGGACCCGTCTTG.
Description
Body ~ 7.04 mm. Head: Antenna 34-segmented. Face width ~ 1.36 × wider than high (1.01:0.74). Width of anterior ocellus as long as POL (010:0.10). Eyes bulged and without interommatidial setae; median width of eye about ~ 1.23 × longer than the median width of gena in lateral view (0.37:0.30). Clypeus 2.20 × longer than its height (0.55:0.25). Galea 1.64 × longer than height of clypeus (0.41:0.25), relatively longer than other members of the genus. Glossa 1.96 × longer than height of clypeus (0.49:0.25). Occipital carina absent. Mesosoma: Notauli weakly impressed at anterior two thirds and absent posteriorly. Scutellar sulcus straight, moderately impressed, medially shallow, laterally relatively deep, without crenula. Pronotum mostly smooth, with ventral longitudinal carina. Metapleuron mostly smooth. Propodeum ~ 0.48 × longer than its median width (0.40:0.83); median longitudinal furrow present, nearly rectangle, anteriorly opened posteriorly closed by nucha, anterior width ~ 0.83 × longer than maximum width (0.05:0.06). Legs: Basal spur on mid tibia ~ 0.60 × longer than length of basitarsus (0.38:0.63). Basal spur on hind tibia ~ 0.54 × longer than length of basitarsus (0.60:1.11). Wings: Fore wing (RS+M)a present; second submarginal cell trapezoid, ~ 2.81 × longer than height (1.32:0.47); 1r absent; 3r absent; 3RSb evenly curved. Hind wing 2r-m absent; 2–1A absent. Metasoma: T1 ~ 2.22 × longer than its posterior width (0.91:0.41), anterior width ~ 0.83 × longer than posterior width (0.34:0.41), dorsally nearly rectangle; Y-shaped suture present. T2 nearly rectangle, ~ 0.49 × longer than its posterior width (0.35:0.71), with straight posterior margin. Hypopygium with median fold. Ovipositor sheath nearly straight, ~ 0.62 × longer than length of hind tibia (1.31:2.11), evenly setose except for base.
Color
Body mostly yellowish pale. The following areas dark: basal antenna (mostly), head, mandible, mesonotum, pronotum, mesopleuron, metapleuron, middle coxa, hind coxa, apical hind tibia, apical hind tarsi, apical metasoma. Fore wing with two bands; stigma mostly pale.
Etymology
Named in honor of Dr. Alexey K. Tishechkin, staff in the Plant Pest Diagnostics Branch, California Department of Food & Agriculture and a former member of the Louisiana State Arthropod Museum at LSU AgCenter, who collected the specimen in Ecuador.
Male
Unknown.
Host
Unknown.
Distribution
Neocardiochiles alexeyi sp. nov. is known from only one female specimen collected from Yasuni Biological Station, Orellana province, Ecuador.
Neocardiochiles braeti , sp. nov.
Material examined
Holotype French Guiana • ♀; Roura, Montagne des chevaux; 22.xi.2008; S.E.A.G.; 2008–2009. Will be deposited in
Diagnosis
Neocardiochiles braeti sp. nov. can be distinguished from other Neocardiochiles species by the combination of the following characters: antenna ≥ 37-segmented; scutellar sulcus weakly impressed (Fig.

Description
Body ~ 8.67 mm. Head: Antenna ≥ 37-segmented, apically broken. Face width ~ 1.46 × longer than its height (1.02:0.70) Width of anterior ocellus ~ 0.96 × longer than POL. Eyes bulged and without interommatidial setae; median width of eye about ~ 0.90 × longer than median width of gena in lateral view (0.44:0.49). Clypeus ~ 2.61 × longer than its height (0.60:0.23). Malar space 0.84 × longer than basal width of mandible. Mesosoma: Notauli anteriorly weakly impressed and absent posteriorly. Scutellar sulcus weakly impressed, medially shallow, laterally relatively deep, without crenula. Pronotum entirely smooth. Propodeum ~ 0.44 × longer than its median width (0.57:1.30); median longitudinal furrow present, elongate isosceles trapezoid, anteriorly opened posteriorly closed by nucha, anterior width ~ 0.33 × longer than maximum width (0.04:0.12). Legs: Basal spur on mid tibia ~ 0.64 × longer than length of basitarsus (0.47:0.74). Basal spur on hind tibia ~ 0.50 × longer than length of basitarsus (0.68:1.37). Wings: Fore wing (RS+M)a vein present; second submarginal cell trapezoid, ~ 3.01 × longer than height (1.66:0.55); 1r absent; 3r absent; 3RSb evenly curved; stigma about ~ 4.42 × longer than wide medially (2.08:0.47). Hind wing 2r-m absent; 2–1A absent. Metasoma: T1 ~ 1.63 × longer than its posterior width (1.19:0.73), anterior width ~ 0.63 × longer than posterior width (0.46:0.73), dorsally isosceles trapezoid; Y-shaped suture partially developed anteriorly, absent posteriorly. T2 nearly trapezoid, ~ 0.42 × longer than its posterior width (0.44:1.06), with curved posterior margin. Hypopygium with median fold. Ovipositor sheath slightly downcurved, ~ 1.03 × longer than length of hind tibia (2.92:2.81), evenly setose except for base.
Color
Head and mesosoma mostly black, metasoma mostly orange and black apically. Legs black or dark brown except for fore tarsus. Fore wing with two bands; stigma pale at apical two thirds.
Etymology
Named in honor of Mr Yves Braet who provided specimens collected in French Guiana for this study.
Male
Unknown.
Host
Unknown.
Distribution
Neocardiochiles braeti sp. nov. is known only from one female specimen collected from Roura, Montagne des chevaux, French Guiana.
Neocardiochiles chriscarltoni , sp. nov.
Material examined
Holotype French Guiana • ♀; Roura, Montagne des chevaux; ix.2009; S.E.A.G.; 2008–2009, Piège Malaise. Will be deposited in
Diagnosis
Neocardiochiles chriscarltoni sp. nov. is most similar to N. braeti sp. nov. However, N. chriscarltoni sp. nov. can be distinguished from other Neocardiochiles species by the combination of the following characters: antenna 38-segmented and with pale apex (Fig.


Molecular data
16S sequences (GenBank accession number: ON059709.1);
CACCTGTTTATCAAAAACATGTCTTTTTGAAAATAATTTAAAGTCCAATCTGCTCAATGATTAATTAATTTAATAGCTGCAATATTTATAATTGTACTAAGGTAGCATAATCATTAGTTTATTAATTGTAAACTTGTATGAACGATTTGATGAAATAAATACTATTTCATTTTAAAAAAATAAATTTTTTTTTTAAGTTAAAAAACTTAAATAATATTAAAAGACGAGAAGACCCTTTAGAATTTTATAATAATAATTTATAAAAATTTTTATATATTTATAAATAATTATTATTTAATTGGGGTGATTATAAAATTTAATAAACTTTTATATAAATAAACAATAATTTTTGAATAAAATATATTTTTTTTAAAAAAATAAATTAAATTACCTAAGGGATAACAGCATAATTTTTTTAAAAAGCACAAATTTATAAAAAAGTTTATGACCTCGATGTTGAATTAAGA.
Description
Body ~ 7.43 mm. Head: Antenna 38-segmented. Face width ~ 1.47 × longer than its height (0.97:0.66). Width of anterior ocellus as long as POL (0.12:0.12). Eyes bulged and without interommatidial setae; median width of eye about ~ 1.32 × longer than the median width of gena in lateral view (0.50:0.38). Clypeus ~ 2.57 × longer than its height (0.59:0.23). Mesosoma: Notauli weakly impressed at anterior half and disappeared posteriorly. Scutellar sulcus weakly impressed, medially shallow, laterally relatively deep, without crenula. Pronotum mostly smooth, with ventral longitudinal carina. Propodeum ~ 0.39 × longer than its median width (0.53:1.36); median longitudinal furrow present, elongate isosceles trapezoid, anteriorly opened posteriorly closed by nucha, anterior width ~ 0.11 × longer than maximum width (0.02:0.18). Legs: Basal spur on mid tibia ~ 0.56 × longer than length of basitarsus (0.38:0.68). Basal spur on hind tibia ~ 0.48 × longer than length of basitarsus (0.52:1.08). Wings: Fore wing (RS+M)a vein present; second submarginal cell trapezoid; 1r absent; 3r absent; 3RSb evenly curved. Hind wing 2r-m absent; 2–1A absent. Metasoma: T1 ~ 1.06 × longer than its posterior width (0.82:0.77), anterior width ~ 0.66 × longer than posterior width (0.51:0.77), dorsally nearly rectangular; Y-shaped suture present. T2 nearly rectangle, ~ 0.30 × longer than its posterior width (0.38:1.28), with curved posterior margin. Hypopygium with median fold. Ovipositor sheath nearly straight and posteriorly enlarged, ~ 0.48 × longer than length of hind tibia (1.07:2.25), evenly setose except for base.
Color
Body mostly melanic; the following areas pale: four apical flagellomeres, basal maxillary and labial palpi, fore tarsi, metanotum (mostly), propodeum, anterior metasoma (mostly). Fore wing with two bands; stigma entirely melanic.
Etymology
Named in honor of Dr Christopher E. Carlton, the emeritus professor in the Department of Entomology at LSU AgCenter.
Male
Unknown.
Host
Unknown.
Distribution
Neocardiochiles chriscarltoni sp. nov. is known only from one female specimen collected from Roura, Montagne des chevaux, French Guiana.
Neocardiochiles fasciipennis , comb. nov.
Material examined
Lectotype Suriname • ♀; Michaelis.
Diagnosis
Morphological characters of Neocardiochiles fasciipennis are only known from holotype. The species is distinguished from other members of Neocardiochiles by the combination of the following characters: body ~ 11 mm; posterior margin of mesopleuron smooth; basal spur on hind tibia slightly shorter than the half length of hind basitarsus (Fig.

Description
See
Male
Unknown.
Host
Unknown.
Distribution
Suriname.
Neocardiochiles franki , sp. nov.
Material examined
Holotype Costa Rica • ♀; 3 km SE of Rio Naranjo, Guanacaste; 1–5.vi.1992; F. D. Parker. Will be deposited in UWIM.
Diagnosis
Neocardiochiles franki sp. nov. is most similar to N. alexeyi sp. nov. However, N. franki sp. nov. can be distinguished from other Neocardiochiles species by the combination of the following characters: antenna ≥ 37-segmented; scutellar sulcus weakly impressed (Fig.

Description
Body ~ 9.36 mm. Head: Antenna ≥ 37-segmented, apically broken. Face width ~ 1.50 × longer than its height (1.11:0.74). Width of anterior ocellus ~ 1.11 × longer than POL (0.10:0.09). Eyes bulged and without interommatidial setae; median width of eye about ~ 1.32 × longer than median width of gena in lateral view (0.47:0.44). Clypeus ~ 2.41 × longer than its height (0.77:0.32). Mesosoma: Notauli weakly impressed at anterior half and absent posteriorly. Scutellar sulcus weakly impressed, medially shallow, laterally relatively deep, without crenula. Pronotum mostly smooth, with ventral longitudinal carina. Propodeum 0.50 × longer than its median width (0.61:1.22); median longitudinal furrow present, elongate isosceles trapezoid, anteriorly opened posteriorly closed by nucha, anterior width ~ 0.18 × longer than maximum width (0.03:0.17). Legs: Basal spur on mid tibia ~ 0.60 × longer than length of basitarsus (0.47:0.78). Basal spur on hind tibia ~ 0.62 × longer than length of basitarsus (0.83:1.33). Wings: Fore wing (RS+M)a vein present; second submarginal cell trapezoid, ~ 2.20 × longer than height (1.81:0.82); 1r absent; 3r absent; 3RSb evenly curved. Hind wing 2r-m absent; 2–1A absent. Metasoma: T1 ~ 1.09 × longer than its posterior width (1.20:1.10), anterior width ~ 0.53 × longer than posterior width (0.58:1.10), dorsally nearly trapezoid; Y-shaped suture present. T2 rectangle, ~ 0.34 × longer than its posterior width (0.54:1.58), with straight posterior margin. Hypopygium with median fold. Ovipositor sheath nearly straight and posteriorly enlarged, ~ 0.58 × longer than length of hind tibia (1.82:3.15), evenly setose except for base.
Color
Body mostly melanic; the following areas pale: maxillary and labial palpi, fore leg, middle femur; mid tibia; basal middle tarsus, hind femur, basal hind tibia, anterior metasoma (mostly). Fore wing with two bands; stigma mostly pale.
Etymology
Named in honor of Dr Frank Parker, the former head of the USDA Bee lab, who collected the specimen.
Male
Unknown.
Host
Unknown.
Distribution
Neocardiochiles franki sp. nov. is known only from one female specimen collected from 3 km SE of Rio Naranjo, Guanacaste, Costa Rica.
Neocardiochiles hasegawai , comb. nov.
Material examined
Non-type specimen Costa Rica • ♀; Guanacaste, Area de Conservación Guanacaste, Sector Santa Rosa, Finca Jenny; 10.86333, -85.57443, 205 m; 14.xii.2010 (host caterpillar collection date); 01.i.2011 (host caterpillar pre-pupal date); 17.i.2011 (parasitoid eclosion date); Johan Vargas; DHJPAR0045389; host caterpillar (10-SRNP-15361; Stenoma cathosiota) on host plant (Roupala montana).
Diagnosis
Neocardiochiles hasegawai is most similar to N. alexeyi sp. nov. and N. franki sp. nov., but the members of N. hasegawai differ from other Neocardiochiles species by possession of the two following characters: mesoscutum entirely pale; hind femur entirely pale.
Molecular data
28S sequences (GenBank accession number: ON040755.1);
ATGAGGAGATTCACTGTTAGCATTACTAGTATTAATGCAAATTATGATATGATTATATATGATTCTTGTGGTCACAATTATTATACTTATTTGTATTATTTTATTGGTTTTGTCAGCATGCACTTCTCCTCTAGTAGAACGTCGCGACCCGTTGAATGTTTATTTATGAGTCACATGGTAGTCTTATGTATTTTATACGCAAGACCAGTGAATTTCTAATAAACTGTTTGACGGTATCTAAAATGGTATTGAGCCGCAAATTTTTTTTTGCGTTAGATTTATCACAAGCTAGGCTTACTTTAAGCAGTACGAATTTTATGTCGTCGTTTAAACTAGTCTGCTGTTAGTGATAATATCTTTAACTGGCTTAATTACCGGTCAGCGATGCTACTGCTTTGGGTACTTACAGGACCCGTCTTG.
Description
See
Male
Body length slightly longer than female (
Host
Reared from caterpillars of Carthara abrupta (Pyralidae) on Roupala montana (Proteaceae) (
Distribution
Costa Rica (ACG).
Neocardiochiles kidonoi , comb. nov.
Material examined
Non-type specimen Costa Rica • ♀; Guanacaste, Area de Conservación Guanacaste, Sector El Hacha, Quebrada Pitahaya; 11.01182, -85.53168; 320 m; 11.ix.2013 (host caterpillar collection date); 14.ix.2013 (host caterpillar pre-pupal date); 02.x.2013 (parasitoid eclosion date); Roster Moraga; DHJPAR0053597; host caterpillar (13-SRNP-22162; Stenoma cathosiota) on host plant (Roupala montana).
Diagnosis
Neocardiochiles kidonoi is most similar to N. victoriae sp. nov., but members of N. kidodoi can be distinguished from other Neocardiochiles members by possession of the following characters: mesoscutum entirely pale; hind femur bicolored; hind tibia mostly pale.
Description
See
Male
Body length slightly shorter than female (
Host
Reared from larvae of Stenoma cathosiota (Depressariidae) on Roupala montana (Proteaceae) (
Distribution
Costa Rica (ACG).
Neocardiochiles victoriae , sp. nov.
Material examined
Holotype French Guiana • ♀; Degrad Laurens, Crique Sapokaï; 95 m; 24.x.–30.x1998; leg. A.E.I. guyane; P Malaise. Will be deposited in
Diagnosis
Neocardiochiles victoriae sp. nov. is most similar to N. kidonoi, but the only knonw female member of N. victoriae differs from other members of Neocardiochiles by the following characters: antenna 40-segmented with entirely reddish flagellomeres (Fig.

Molecular data
28S sequences (GenBank accession number: ON040754.1);
ATGAGGAGATTCACTGTTAGCATTACTAATATTAATGCAAATTATGATATCATTATATGATCCTTGTGGTCACAATTATTATACTTATTTGTATTATTTTATTGGTTTTGTCAGCATGCACTTCTCCTCTAGTAGAACGTCGCGACCCGTTAAATGTTTATTTATGAGTCACATGGTAGTCTTATGTATTTTATACGCAAGACCAGTGAATTTCTAATAAACTGTTTGACGGTATCTAAAATGGTATTGAGCCGCAAATTTTTTTTGCGTTAGATTTATTACAAGCTAGGCTTACTTTAAGCAGTACGAATTTTATGTCGTCGTTTAAACTAGTCTGCTGTTAGTGATAATATCTTTAACTGGCTTAATTACCGGTCAGCGATGCTACTGCTTTGGGTACTTACAGGACCCGTCTTG.
16S sequences (GenBank accession number: ON059710.1);
TCCCCTGTTTATCAAAAACATGTCTTATTGAAAATAATTTTAAGTCAAATCTGCTCAATGATAATTTTATTAAATAGCTGCAGTAAATATGACTGTACTAAGGTAGCATAATAAATAGTTTATTAATTATAAACTTGTATGAAAGATTTAATGTAATAAATACTGTTTCAATTTAAAAAAATAAATTTTTTTTTTAAGTAAAAAAACTTAAATAAAATTAAAAGACGAGAAGACCCTATAGAATTTTATAAATTAATTTTAATTAATTTATTTTTAATTAAAATTAAATTATTTAATTGGGGAGATTATAAAATTTAAAAAACTTTTATATAAATTTACAATAATTATTGAATAAAATATAATTTTTAAAAAAAATAAAATAAATTACCTTAGGGATAACAGCATAATTTTTTTTTTAAGTTCGTATTACTAAAAAAGATTATGACCTCGATGTTGAATTAAGA
Description
Body ~ 9.35 mm. Head: Antenna 40-segmented. Face width ~ 1.73 × longer than its height (1.28:0.74). Width of anterior ocellus ~ 1.15 × longer than POL (0.15:0.13). Eyes bulged and without interommatidial setae; median width of eye about ~ 1.04 × longer than median width of gena in lateral view (0.58:0.56). Clypeus ~ 2.39 × longer than its height (0.74:0.31). Mesosoma: Notauli weakly impressed at anterior half and absent posteriorly. Scutellar sulcus weakly impressed, medially shallow, laterally relatively deep, without crenula. Pronotum mostly smooth, with ventral longitudinal carina. Metapleuron mostly smooth. Propodeum ~ 0.39 × longer than its median width (0.66:1.68); median longitudinal furrow present, elongate isosceles trapezoid, anteriorly opened posteriorly closed by nucha, anterior width ~ 0.19 × longer than maximum width (0.03:0.16). Legs: Basal spur on mid tibia ~ 0.71 × longer than length of basitarsus (0.62:0.87). Basal spur on hind tibia 0.50 × longer than length of basitarsus (0.84:1.68); claw pectinate. Wings: Fore wing (RS+M)a vein present; second submarginal cell trapezoid; 1r absent; 3r absent; 3RSb evenly curved. Hind wing 2r-m absent; 2–1A absent. Metasoma: T1 ~ 1.39 × longer than its posterior width (1.42:1.02), anterior width ~ 0.56 × longer than posterior width (0.57:1.02), dorsally isosceles trapezoid; Y-shaped suture present. T2 nearly oval, ~ 0.39 × longer than its posterior width (0.49:1.27), with curved posterior margin. Hypopygium with median fold. Ovipositor sheath slightly downcurved, ~ 1.04 × longer than length of hind tibia (3.38:3.25), evenly setose except for base.
Color
Body mostly orange. The following areas dark: antenna (reddish brown), head, ventral fore notum, fore coxa, fore trochanter and trochantellus (mostly), fore femur (mostly); middle coxa, middle trochanter and trochantellus (mostly), middle femur (mostly), mid tibia (apically); hind femur (apically); hind tibia (except for inner tibia), fifth to last terga (dorsally), ovipositor sheath. Fore wing with two bands; stigma mostly pale.
Etymology
Named in honor of Ms Victoria Bayless, a curator in the Louisiana State Arthropod Museum at LSU AgCenter and a former president of the Coleopterists Society, who is the best friend in LSU and has red hair as the specimen has reddish antennae.
Male
Unknown.
Host
Unknown.
Distribution
Neocardiochiles victoriae sp. nov. is known only from one female specimen collected from Degrad Laurens, Crique Sapokaï, French Guiana.
Neocardiochiles whitfieldi , comb. nov.
Material examined
Holotype Mexico • ♂; 16 km north of Autlan, Jalisco; 7.vii.1984; Carrol, Schaffner, Friedlander. Deposited in
Diagnosis
The known male of Neocardiochiles whitfieldi can be easily distinguished from other members of Neocardiochiles by the following color characters: head and mesosoma darker; wings entirely melanic; metasoma orange.
Description
(Male). See
Female
Unknown.
Host
Unknown.
Distribution
Mexico.
Acknowledgements
We thank Drs Daniel Janzen and Winnie Hallwachs at the University of Pennsylvania, Dr Michael Sharkey at HIC, Dr Scott Shaw at UWIM, Dr Karen Wright at
References
- Belshaw R, Lopez-Vaamonde C, Degerli N, Quicke DL (2001) Paraphyletic taxa and taxonomic chaining: evaluating the classification of braconine wasps (Hymenoptera:Braconidae) using 28S D2–3 rDNA sequences and morphological characters. Biological Journal of the Linnean Society 73: 411–424. https://doi.org/10.1111/j.1095-8312.2001.tb01370.x
- Belshaw R, Quicke DL (1997) A molecular phylogeny of the Aphidiinae (Hymenoptera: Braconidae). Molecular Phylogenetics Evolution 7(3): 281–293. https://doi.org/10.1006/mpev.1996.0400
- Brullé A (1846) Histoire Naturelles des Insects: Hymenopteres. Tome quatrieme. Librairie Encyclopedique de Roret, Paris, 689 pp. https://doi.org/10.5962/bhl.title.9005
- Campbell B, Heraty J, Rasplus JY, Chan K, Steffen-Campbell J, Babcock C (2000) Molecular systematics of the Chalcidoidea using 28S-D2 rDNA. Hymenoptera evolution, biodiversity and biological control. CSIRO Publishing, Collingwood, Australia 26: 59–73.
- Chen YI, Pike KS, Greenstone MH, Shufran KA (2006) Molecular markers for identification of the hyperparasitoids Dendrocerus carpenteri and Alloxysta xanthopsis in Lysiphlebus testaceipes parasitizing cereal aphids. Biocontrol 51(2): 183–194. https://doi.org/10.1007/s10526-005-1518-0
- Dabek EZ, Whitfield JB, Hallwachs W, Janzen DH (2020) Two new reared species of Heteropteron Brullé (Hymenoptera, Braconidae, Cardiochilinae) from northwest Costa Rica, with the first definitive host records for the genus. Journal of Hymenoptera Research 77: 151–165. https://doi.org/10.3897/jhr.77.50577
- Dallwitz MJ, Paine TA, Zurcher EJ (1999) User’s guide to the DELTA Editor. http://delta-intkey.com [accessed on Mar. 12. 2022]
- Dallwitz MJ, Paine TA, Zurcher EJ (2000) Principles of interactive keys. http://delta-intkey.com [accessed on Mar. 12. 2022]
- Dangerfield PC, Austin AD, Whitfield JB (1999) Systematics of the world genera of Cardiochilinae (Hymenoptera: Braconidae). Invertebrate Systematics 13(6): 917–976. https://doi.org/10.1071/IT98020
- Darriba D, Posada D, Kozlov AM, Stamatakis A, Morel B, Flouri T (2020) ModelTest-NG: a new and scalable tool for the selection of DNA and protein evolutionary models. Molecular Biology and Evolution 37(1): 291–294. https://doi.org/10.1093/molbev/msz189
- Dowton M, Austin AD (1994) Molecular phylogeny of the insect order Hymenoptera: apocritan relationships. Proceedings of the National Academy of Sciences of the United States of America 91(21): 9911–9915. https://doi.org/10.1073/pnas.91.21.9911
- Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research 32(5): 1792–1797. https://doi.org/10.1093/nar/gkh340
- Edler D, Klein J, Antonelli A, Silvestro D (2021) raxmlGUI 2.0: a graphical interface and toolkit for phylogenetic analyses using RAxML. Methods in Ecology and Evolution 12(2): 373–377. https://doi.org/10.1111/2041-210X.13512
- Fagan-Jeffries EP, Austin AD (2020) Synopsis of the parasitoid wasp genus Cotesia Cameron, 1891 (Hymenoptera: Braconidae: Microgastrinae) in Australia, with the description of seven new species. European Journal of Taxonomy 667: 1–70. https://doi.org/10.5852/ejt.2020.667
- Fernandez-Triana J, Whitfield J, Rodriguez J, Smith M, Janzen D, Hajibabaei M, Burns J, Solis A, Brown J, Cardinal S, Goulet H, Hebert P (2014) Review of Apanteles sensu stricto (Hymenoptera, Braconidae, Microgastrinae) from Area de Conservación Guanacaste, northwestern Costa Rica, with keys to all described species from Mesoamerica. ZooKeys 383: 1–565. https://doi.org/10.3897/zookeys.383.6418
- Fernandez-Triana J, Boudreault C, Dapkey T, Smith MA, Hallwachs W, Janzen D (2019) A revision of Dolichogenidea (Hymenoptera, Braconidae, Microgastrinae) with the second mediotergite broadly rectangular from Area de Conservación Guanacaste, Costa Rica. ZooKeys 835: 87–123. https://doi.org/10.3897/zookeys.835.33440
- Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3: 294–299.
- Harris RA (1979) Glossary of surface sculpturing. Occasional Papers in Entomology 28: 1–31. https://doi.org/10.5281/zenodo.26215
- Hasegawa M, Kishino H, Yano T (1985) Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. Journal of Molecular Evolution 22(2): 160–174. https://doi.org/10.1007/BF02101694
- Hebert PDN, Penton EH, Burns JM, Janzen DH, Hallwachs W (2004) Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proceedings of the National Academy of Sciences of the United States of America 101(41): 14812–14817. https://doi.org/10.1073/pnas.0406166101
- Kang I, Chapman EG, Janzen DH, Hallwachs W, Dapkey T, Smith MA, Sharkey MJ (2017) Revision of the species of Lytopylus from Area de Conservación Guanacaste, northwestern Costa Rica (Hymenoptera, Braconidae, Agathidinae). ZooKeys 721: 93–158. https://doi.org/10.3897/zookeys.721.20287
- Maddison WP, Maddison DR (2021) Mesquite: a modular system for evolutionary analysis. Version 3.70. http://www.mesquiteproject.org [accessed on Oct. 10. 2021]
- Madeira F, mi Park Y, Lee J, Buso N, Gur T, Madhusoodanan N, Basutkar P, Tivey ARN, Potter SC, Finn RD, Lopez R (2019) The EMBL-EBI Search and Sequence Analysis Tools APIs in 2019. Nucleic Acids Research 47: W636–W641. https://doi.org/10.1093/nar/gkz268
- Meierotto S, Sharkey MJ, Janzen DH, Hallwachs W, Hebert PDN, Chapman EG, Smith MA (2019) A revolutionary protocol to describe understudied hyperdiverse taxa and overcome the taxonomic impediment. Deutsche Entomologische Zeitschrift 66(2): 119–145. https://doi.org/10.3897/dez.66.34683
- Mercado I, Wharton RA (2003) Mexican cardiochiline genera (Hymenoptera: Braconidae), including a preliminary assessment of species-groups in Toxoneuron Say and Retusigaster Dangerfield, Austin and Whitfield. Journal of Natural History 37(7): 845–902. https://doi.org/10.1080/00222930110097167
- Murphy N, Banks JC, Whitfield JB, Austin AD (2008) Phylogeny of the microgastroid complex of subfamilies of braconid parasitoid wasps (Hymenoptera) based on sequence data from seven genes, with an improved estimate of the time of origin of the lineage. Molecular Phylogenetics and Evolution 47: 378–395. https://doi.org/10.1016/j.ympev.2008.01.022
- Nei M, Kumar S (2000) Molecular Evolution and Phylogenetics. Oxford University Press, Oxford, 352 pp.
- Papp J (2014) A revisional study on Szépligeti’s cardiochiline type specimens deposited in the Hungarian Natural History Museum, Budapest (Hymenoptera, Braconidae: Cardiochilinae). In Annales historico-naturales Musei nationalis hungarici 106: 169–214.
- Sharkey MJ, Wharton RA (1997) Morphology and terminology. In: Wharton RA, Marsh PM, Sharkey MJ (Eds) Manual of the New World genera of the family Braconidae (Hymenoptera). Special Publication of the International Society of Hymenopterists, No 1, Washington DC, 19–37.
- Sharkey MJ, Janzen DH, Hallwachs W, Chapman EG, Smith MA, Dapkey T, Brown A, Ratnasingham S, Naik S, Manjunath R, Perez K, Milton M, Hebert P, Shaw SR, Kittel RN, Solis MA, Metz MA, Goldstein PZ, Brown JW, Quicke DLJ, van Achterberg C, Brown BV, Burns JM (2021a) Minimalist revision and description of 403 new species in 11 subfamilies of Costa Rican braconid parasitoid wasps, including host records for 219 species. ZooKeys 1013: 1–665. https://doi.org/10.3897/zookeys.1013.55600
- Sharkey MJ, Baker A, McCluskey K, Smith A, Naik S, Ratnasingham S, Manjunath R, Perez K, Sones J, D’Souza M, Jacques BS, Hebert P, Hallwachs W, Janzen D (2021b) Addendum to a minimalist revision of Costa Rican Braconidae: 28 new species and 23 host records. ZooKeys 1075: 77–136. https://doi.org/10.3897/zookeys.1075.72197
- Slater‐Baker MR, Austin AD, Whitfield JB, Fagan‐Jeffries EP (2022) First record of miracine parasitoid wasps (Hymenoptera: Braconidae) from Australia: molecular phylogenetics and morphology reveal multiple new species. Austral Entomology 61(1): 49–67. https://doi.org/10.1111/aen.12582
- Smith MA, Rodriguez JJ, Whitfield JB, Deans AR, Janzen DH, Hallwachs W, Hebert PD (2008) Extreme diversity of tropical parasitoid wasps exposed by iterative integration of natural history, DNA barcoding, morphology, and collections. Proceedings of the National Academy of Sciences 105(34): 12359–12364. https://doi.org/10.1073/pnas.0805319105
- Smith MA, Fernández-Triana JL, Eveleigh E, Gómez J, Guclu C, Hallwachs W, Hebert PDN, Hrcek J, Huber JT, Janzen DH, Mason PG, Miller S, Quicke DLJ, Rodriguez JJ, Rougerie R, Shaw MR, Várkonyi G, Ward D, Whitfield JB, Zaldivar-Riveron A (2012) DNA barcoding and the taxonomy of Microgastrinae wasps (Hymenoptera, Braconidae): impacts after 8 years and nearly 20,000 sequences. Molecular Ecology Resources 13(2): 168–176. https://doi.org/10.1111/1755-0998.12038
- Szépligeti V (1908) Braconiden aus der Sammlung des Ungarischen National-Museums. II Teil. Annales Historico-Naturales Musei Nationalis Hungarici 6: 397–427.
- Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution 38(7): 3022–3027. https://doi.org/10.1093/molbev/msab120
- Whitfield JB, Dangerfield PC (1997) Subfamily Cardiochilinae. In: Wharton RA, Marsh PM, Sharkey MJ (Eds) Manual of the New World Genera of the Family Braconidae (Hymenoptera). International Society of Hymenopterists, Washington DC, 177–183.
- Yu DS, van Achterberg CV, Horstmann K (2016) Taxapad 2016, Ichneumonoidea 2015. Database on flash-drive. Ottawa, Ontario.
Supplementary materials
Maximum likelihood (ML) phylogeny based on 16S data Maximum likelihood (ML) phylogeny based on 16S data indicating the relationships of Neocardiochiles chriscarltoni sp. nov. and N. victoriae sp. nov. Protomicroplitis calliptera (Hymenoptera: Braconidae: Microgastrinae) is included as the outgroup.
Data type: Image
Explanation note: Maximum likelihood (ML) phylogeny based on 16S data indicating the relationships of Neocardiochiles chriscarltoni sp. nov. and N. vitoriae sp. nov. Protomicroplitis calliptera (Hymenoptera: Braconidae: Microgastrinae) is included as the outgroup.
Maximum likelihood (ML) phylogeny based on 28S data Maximum likelihood (ML) phylogeny based on 28S data indicating the relationships of Neocardiochiles chriscarltoni sp. nov. and N. victoriae sp. nov. Protomicroplitis calliptera (Hymenoptera: Braconidae: Microgastrinae) is included as the outgroup.
Data type: Image
Explanation note: Maximum likelihood (ML) phylogeny based on 28S data indicating the relationships of Neocardiochiles chriscarltoni sp. nov. and N. vitoriae sp. nov. Protomicroplitis calliptera (Hymenoptera: Braconidae: Microgastrinae) is included as the outgroup.
Neocardiochiles_DELTA_Key
Data type: Delta interactive key
Explanation note: Files and sources of DELTA Interactive Key.
Character matrix for Heteropteron, Wesmaelella, and Neocardiochiles (Hymenoptera: Braconidae: Cardiochilinae)
Data type: Nexus File
Explanation note: Character matrix for Heteropteron, Wesmaelella, and Neocardiochiles (Hymenoptera: Braconidae: Cardiochilinae) used in phylogenetic analysis.